
Изменчивость, информация, и сотворенный род
Автор: Карл Виланд (англ. Carl Wieland)
Источник: creation.com
Перевод: Владимир Матвеев
Редактура: Владимир Силенок
Резюме
Все наблюдаемые биологические изменения имеют в своей основе только сохранение или разрушение генетической информации. Поэтому мы не наблюдаем той эволюции которую обычно имеют ввиду под словом «эволюция». По соображениям логики, практичности и стратегии в статье предлагается:
- Избегать использования термина «микроэволюция».
- Переосмыслить наше употребление всей концепции изменчивости внутри «библейского рода».
- Отказаться от использования таксономических определений библейского рода кроме аксиомных.
Наиболее часто в популярной литературе по эволюции встречается идея о том, что если наблюдаемые сейчас небольшие изменения в последовательных поколениях организмов перенести назад во времени, то этим можно объяснить всю эволюцию от одной клетки до человека. Креационисты же, якобы, провели воображаемую черту, так сказать, «Линию Мажино», за пределами которой изменчивость не дозволена, а так называемая микро-эволюция или изменчивость «по роду их» не выходит за эту границу. В конце концов, если креационист так говорит, то он имеет ввиду, что комары не превращаются в слонов или мотыльков, потому что это — чрезмерное упрощение. С другой стороны, неодарвинист может справедливо заметить, что он и не ожидает увидеть такого рода изменений в период одной человеческой жизни. Пост-неодарвинист может заявить, что наша выборка из геологического времени слишом мала чтобы ожидать появления «некого монстра» или вообще любого другого существенного изменения.
Другой причиной, по которой позиция креациониста часто выглядит слабой, является тот факт, что о нас думают как о людях признающих биологическую изменчивость только потому что мы вынуждены делать это на основе наблюдений, но при этом избегаем обсуждать потенциальные последствия изменчивости, используя отговорку, что изменчивость не заходит достаточно далеко. Мы выглядим как люди передвигающие «Линию Мажино» в зависимости от степени наблюдаемой изменчивости. Однако, далее в этой статье будет показано, что такой подход ложен и является лишь карикатурой на позицию креационизма, и что пределы изменчивости, на самом деле, определяются на уровне генетической информации.
СОТВОРЕННЫЕ РОДЫ
Наблюдаемая изменчивость очевидно имеет свои пределы. Возникает соблазн использовать этот факт для демонстрации существования сотворенных родов, и изменчивости, которая ограничена такими родами.
Однако, такая аргументация является круговой и потому она уязвима. Поскольку креационисты по определению считают всю изменчивость ограниченной «пределами сотворенного рода», (см., например, такую позицию у Американского общества исследователей креационизма), как тогда нам доказать с помощью наблюдений что изменчивость ограничена пределами сотворенного рода? Иначе говоря, мы никогда не наблюдали изменчивость «внутри рода», поскольку всякий раз когда две разновидности происходят из одного общего источника, их считают одним и тем же родом. Не удивительно, поэтому, что эволюционисты требуют от нас точного определения термина «сотворенный род» поскольку только в этом случае наше заявление что «изменчивость ограничена только сотворенным родом» перестает быть тавтологией и проверяема научно. Однако, круговое обоснование не обесценивает концепцию сотворенных родов. Подобным же образом в определении естественного отбора используется круговое обоснование (выживают наиболее приспособленные, а наиболее приспособленные это те которые выжили), но тем не менее оно является логичной концепцией, которую легко проверить. Мы всего лишь хотим сказать, что аргументы являющиеся по своей природе круговыми нельзя использовать как независимое доказательство сотворенных родов.
Когда я говорю, что такое независимое доказательство может быть невозможным по природе самих вещей, мое заявление ни коим образом не является оправданием. Например, давайте предположим что мы обнаружили остатки взорвавшегося острова- камни, деревья, песок и т.д. Восстановить первоначальное взаимное расположение этих элементов может оказаться в принципе невозможным. Однако, это не означает, что нельзя сделать вывода, с большой степенью уверенности, что настоящее расположение остатков взрыва согласуется со взрывом, описанным очевидцем, и не вызвано другим каким-то механизмом.
Подобным же образом, мы можем показать, что наблюдения в природе хорошо согласуются с библейской концепцией сотворенных родов и не согласуются с идеей эволюции. Это лучше всего сделать если обратить внимание на генетически-информационную основу всякого биологического изменения. Такой подход более реалистичен и обнаруживает больше, чем уровень или степень морфологического изменения.
Суть проблемы состоит в качестве, а не в количестве. Дело не в том, что поезду не хватило времени проехать достаточно далеко, дело в том что он поехал в неправильном направлении. Пределы изменчивости- наблюдаемые или ненаблюдаемые- неизбежно проявят себя, когда в генофондах будет исчерпана «функционально эффективная» («телеономическая») генетическая информация. Полное понимание этого устраняет представление о креационисте, который крутит педали назад и перечерчивает линию последнего противостояния в ответ на новые наблюдения новых разновидностей организмов.
Этот факт также делает ненужным разделение на «микро» и «макроэволюцию. Я считаю, что креацианистам луше вообще избегать этих терминов, которые только порождают путаницу и вводят в зблуждение. Слово «эволюция» обычно означает некоторое изменение, которое, в конечном счете, превратит простейшее в человека или рептилию в птицу и т. д. Я надеюсь показать, что в рамках этого понимания, мы вообще не увидим никакой эволюции. Когда мы говорим : «я принимаю микроэволюцию, но не макроэволюцию» мы усиливаем представление о том, что речь идет о количественных изменениях, что неправильно. Речь идет о разных типах изменений.
Это не просто мелкая семантика, но это предмет, имеющий психологическое и тактическое значение. Конечно, кто-то может сказать что «микроэволюция» имеет смысл, когда этот термин каким-то образом определен, но последствия употребления этого слова, значение, которое оно содержит и влияние, которое оно оказывает таковы, что делать уступку было бы неразумно. Микроэволюция как изменение, каким бы малым оно ни было, которое однозначно привело бы к увеличению информации, никогда в природе не наблюдалась.
Так или иначе, ведущие биологи теперь сами признают, что «макроэволюция» это не есть «микроэволюция» (используя их же терминологию), отличие которой состоит только в большей протяженности времени. В ноябре 1980 года, в Чикагском полевом музее естественной истории состоялась конференция нескольких самых известных мировых лидеров эволюционной биологии, которая была посвящена «Макроэволюции». Поместив сообщение о конференции в журнале Science, Роджер Левин написал:
«Центральный вопрос чикагской конференции состоял в том, чтобы решить, можно ли механизмы лежащие в основе микроэволюции экстраполировать для объяснения макроэволюции. Несмотря на риск нанесения ущерба точке зрения некоторых участников конференции, можно определенно ответить Нет.»[1]
В одной из ссылок на слова Франциско Айала (профессора генетики Калифорнийского университета) было упомянуто, что:
«… принимая во внимание современные высказывания палеонтологов, я уверен, что мелкие изменения не накапливаются.»[2]
Учитывая тот факт, что данная статья, по сути, приходит к такому же выводу, ее вряд ли можно назвать радикальной. Несмотря на это, большое число даже хорошо образованных людей остается в неведении. Иначе говоря, они думают, что верна формула: «(Большое изменение) = (Маленькие изменения) Х (Миллионы лет)».
КОНЦЕПЦИЯ ИНФОРМАЦИИ
Буквы на этой странице, т.е. вещество чернил и бумаги, подчиняются законам физики и химии, но эти законы не определяют информации, которая в них записана. Информация может зависеть от материального носителя, который был использован для ее хранения, передачи и поиска, но она не является его свойством. Например, идеи изложенные в данной статье сначала родились в уме, а потом были перенесены на материальный носитель. Биологические молекулы живых существ несут гигантские объемы информации, но опять-таки информация не является их химическим свойством. Как таковая, информация не является частью материи и не подчиняется физическим законам. Она возникает из порядка, из способа построения букв генетического «алфавита» клетки. Этот порядок сборки молекул должен быть привнесен извне, и не является изначальным свойством самих молекул. Живые организмы передают эту информацию из поколения в поколение. Характер последовательности оснований в молекуле ДНК отражает подробный план, определяющий все свойства организма. В конечном счете, наследуемые биологические изменения являются прямым следствием изменений в этой информации. Гены можно рассматривать как «предложения» генетической информации записанной на языке ДНК.
Теперь представьте себе первую популяцию организмов, живущих на «примитивной земле» эволюциониста. Эта, так называемая, клетка конечно же будет содержать в себе массу генетической информации, которой однако будет гораздо меньше, чем в генофонде ее сегодняшних потомков, например, в генофонде людей. Эволюционист предполагает, что эта «телеграмма» явилась родоначальницей целых «энциклопедий», содержащих сведения, которые имеют смысл и записаны в форме полезных генетических предложений. (Ниже поясняется, что означают термины «имеющий смысл» и «полезность» в биологическом контексте). Итак он должен объяснить происхождение во времени новых смысловых предложений. В конечном счете их единственным источником мог быть только процесс мутирования.[3]
Возвращаясь к аналогии напечатанной страницы, информация, записанная в генах живого организма копируется во время размножения таким же образом как это делает пишущая машинка повторяя процесс снова и снова. Мутация- это случайное нарушение, «ошибка копирования». Хотя считается что большинство таких ошибок вредны и бессмысленны, эволюционисты предполагают, что в определенных условиях среды они могут оказаться полезными, и тогда организм обладающий такой мутацией имеет больше шансов выжить и размножиться. Теперь мы посмотрим на информационную основу других механизмов биологической изменчивости. Мы увидим, почему они не порождают новых предложений и в результате, почему эволюционист привлекает для своих объяснений мутации того или иного сорта.
1. Менделевская изменчивость
Это такой механизм, который отвечает за большинство новых разновидностей организмов, возникающих в результате экспериментов по скрещиванию, а также это то, что мы наблюдаем в природе и о чем делаем выводы. Половое размножение позволяет комбинировать пакеты информации многими разными способами, но не производит новых пакетов или предложений. Например, когда многочисленные породы собак были выведены из «дикого типа», то это достигалось через отбор желаемых признаков в последовательных поколениях, таким образом, что гены или фразы-предложения, соответствующие этим признакам изолировались в определенных линиях. Хотя некоторые из этих предложений нельзя было увидеть в первоначальной популяции, они, тем не менее, в ней уже присутствовали. (На время мы забудем мутационный процесс, поскольку подобные новые разновидности могут появляться в генофонде независимо от новых мутаций. Некоторые собаки безусловно обладают признаками мутаций.)
Такой тип изменчивости может наблюдаться только если уже существует некое хранилище таких предложений из которых можно выбирать. Естественный (или искуственный) отбор может объяснить выживание наиболее приспособленных, но не приток наиболее приспособленных приходящих извне — вот в чем состоит настоящий вопрос. Такая менделевская изменчивость ничего не говорит нам о том, откуда возникла генетическая информация в популяции, которая уже существует. Поэтому для того чтобы продемонстрировать эволюцию, движущуюся «вверх», требуются изменения другого сорта, потому что добавления новых полезных «предложений» не произошло. И это наблюдается несмотря на тот факт, что появляются не только новые разновидности, но даже новые виды. Если определить вид как свободно скрещивающуюся природную единицу, легко увидеть, что новые виды могут появляться без направленных «вверх» изменений. То есть без добавления нового кодирования информации, требуемой для новых функций. Например, мутация может вызвать дефект, ведущий к появлению генетического барьера или больших физических различий, подобных тем, которые наблюдаются в размерах собак, принадлежащих к породам датский дог и чихуахуа, делающим невозможным их скрещивание в природе.
Немного удивляет тот факт, что до сих пор мы по-прежнему встречаем пристрастие некоторых креационистов к концепции, согласно которой новые виды никогда не появлялись. Даже если кто-то и предложил бы считать, что это и так, все-таки в полевых наблюдениях можно найти «ряды» организмов, которые почти определенно указывают на то, что два репродуктивно-изолированных вида возникли из одного и того же генофонда предков. Те же самые креационисты, которые не одобряют подобного взгляда, наверное охотно согласились бы, что различные виды принадлежащие к типу «собака», включая наверное волков, лис, шакалов, койотов и домашних собак, произошли от одного общего предка. Поэтому, зачем нам отрицать возможность такого явления в наблюдениях настоящего времени? В этом нет необходимости ни с билейской, ни с научной точки зрения. Более того, когда определенно будет найдено появление нового вида, это лишний раз даст повод критиковать креационистов.
Когда мы видим новые разновидности организмов возникающие в результате искуственного отбора или скрещивания, то это ни что иное как истощение уже имевшейся информации родительской популяции и уменьшение генетического потенциала пригодного для новой изменчивости. Если вы попытаетесь вывести собачку чихуахуа из популяции датского дога или наоборот, то вы обнаружите, что ваша популяция не имеет необходимых «грамматических предложений». Такая ситуация возникает в результате потери части исходного генофонда, забракованной в процессе селекции.
Таким образом, то что поначалу казалось прекрасным примером измений, ведущих к появлению новых признаков, при внимательном рассмотрении с позиций генетики, обернулось потерей информации и движением вниз. Число информационных предложений, носимых каждой подгруппой уменьшилось, что сделало популяцию потенциально менее приспособляемой в свете будущих изменений среды. Экстраполяция такого процесса в будущее время не приведет к эволюции вверх, но, в конечном счете, вызовет вымирание организмов и развитие дефицита генетической информации в популяциях.
2. Полиплоидия
В этом случае, опять-таки, не появятся новые информационные предложения сверх уже существующих. Полиплоидия- это всего лишь умножение («фотокопирование») уже имеющейся информации.
3. Гибридизация
Снова, мы не видим новых информационных предложений. Здесь имеет место лишь смешивание двух уже присутствующих наборов информации.
4. Мутирование
Поскольку мутации, по сути, есть случайные нарушения, не удивительно, что в большинстве случаев они вредны, летальны или не несут полезной информации для функционирования и выживания организма. Случайные нарушения сложного и упорядоченного генетического кода привносят «шум» и хаос, а не новое смысловое содержание, не новую функцию или более сложное устройство, которое, на самом деле, обычно утрачивается. Однако, можно представить себе, что иногда в этом сложном мире «разрушительное» изменение может принести некоторую пользу. Например, если мы выкинем из генома некое кодирующее предложение, то это может привести к полезному укорачиванию длины ног у овец (а такая мутация действительно существует), благодаря которой, овцы перестанут перепрыгивать через забор и убегать с фермы. Или если жук, живущий на изолированном острове, где дуют сильные ветра будет подвержен мутации, вызывающей потерю или повреждение информации, кодирующей развитие крыльев, тогда его бескрылые потомки не будут уносимы в море и получат селективное преимущество. Слепые рыбы в пещерах, и устойчивость к антибиотикам – такие же примеры небольшого числа «полезных» мутаций, которые не ведут к увеличению функциональной сложности, как того требует теория эволюции. От случайных изменений не ожидают увеличения функциональной сложности.
Теперь мы подошли к моменту, когда некоторые станут возражать, говоря что мы злоупотребляем терминами «полезный», «имеющий смысл», «функциональный» и т.д. Они скажут, что если некоторое изменение увеличивает вероятность выживания, тогда по определению, оно имеет биологический «смысл» и является полезным. Но такой подход подразумевает, что живые системы ничем кроме выживания не занимаются, в то время как они и их субсистемы имеют специфические функции и реализуют проекты. Иными словами, они несут телеономическую информацию. В этом состоит одно из главнейших различий между живыми и неживыми объектами (исключая машины). Проекты живых систем не всегда ведут к преимуществу в размножении или выживании- на самом деле, они могут не иметь особого отношения к выживанию, тем не менее они выполняются очень эффективно. Конечно, как всегда делается дарвиновское допущение, о том что на определенном этапе эволюционной истории организма данный проект имел репродуктивную ценность и преимущество в размножении. (Например, рыба-брызгун, с ее поразительным умением сбивать насекомых струей, которое ей не обязательно иметь для выживания в настоящее время.) Однако, поскольку эти допущения не поддаются проверке, было бы законно говорить о генетической информации в телеономическом смысле и независимо от любого потенциального преимущества для выживания.
Сегодняшние генофонды несут в себе огромные количества закодированной информации предписывающей исполнение определенных проектов и функций, которые отсутствуют в теоретической «первоначальной» клетке. Следовательно для того чтобы обосновать возможность эволюции от простейшего до человека, необходимо представить примеры, когда мутации добавляли новое «информационное предложение» или новый генный код, обеспечивающий выполнение нового проекта или функции.
В настоящее время, нам не известно ни одной мутации, которая бы привела к такому возрастанию функциональной сложности. Соображения теории вероятности похоже также устраняют такую возможность, или, по крайней мере, делают такое событие крайне маловероятным, слишком маловероятным чтобы спасти теорию эволюции даже в масштабе времени многих миллиардов лет.
Для дополнительной иллюстрации возмем молекулу гемоглобина человека. Она выполняет проект переноса и доставки кислорода в красных кровяных тельцах весьма эффективным образом. Существует ген или «информационное предложение», которое кодирует производство гемоглобина. Известна мутация (точнее три отдельных мутации, которые дают один и тот же результат), при которой происходит замена одной единственной буквы в предложении. Если вы унаследуете такое изменение от обоих родителей, вы серьезно заболеете, болезнью называемой серповидно-клеточной анемией и при этом долго вы не проживете. Несмотря на это, эволюционисты часто используют этот пример для подтверждения существования «выгодных мутаций». Это потому что если вы унаследуете эту мутацию только от одного родителя, ваши красные кровяные тельца тоже будут подвержены изменениям, но не достаточно сильным чтобы повлиять на ваше выживание. Между тем они не дадут малярийным паразитам эффективно использовать их в качестве клеток-хозяев. В результате, у вас возникнет некоторый иммунитет к малярии, что повысит успех выживания в малярийных областях. Этот пример показывает нам как функционально эффективная молекула гемоглобина превратилась в функционально-поврежденную молекулу гемоглобина. Ген, поврежденный мутацией, вызвавшей эту болезнь, поддерживается на высоком уровне в районах с эндемичной малярией благодаря феномену случайного гетерозиготного превосходства. Его повреждающий эффект для части потомства уравновешивается эффектом защищающим от малярии. Это безусловно не является изменением направленным «вверх». Мы не увидели эволюции эффективного механизма переноса кислорода. Мы не увидели улучшения механизма переноса гемоглобином.
Еще одна поверхностная, но, вероятно, полезная аналогия. Давайте допустим, что некий секретный агент посылает ежедневные телеграммы с территории врага. В тексте говорится: «сегодня враг не готовится к атаке». Но вот однажды при пересылке возникает ошибка, в результате которой слово «не» теряется. Очень вероятно что такая ошибка нанесет большой вред, вплоть до возникновения ядерной войны. Но возможно также что в необычной ситуации она может обернуться пользой (например, во время тестирования отказоустойчивых механизмов). Однако, это не означает, что именно такое изменение потребуется чтобы начать превращение телеграммы в энциклопедию.
Крайне малое число «выгодных» мутаций наблюдавшихся в природе, по-просту, не тот тип изменений, который требуется для эволюции – мы не видим добавления новых предложений которые несут в себе смысл и информацию. Опять-таки, к удивлению, можно часто прочесть работы креационистов, которые настаивают, что выгодные мутации не встречаются. Если выгодность определить только как способность выживать, тогда мы не будем ожидать этого во всех случаях, и по сути, так оно и есть — «выгодные» мутации только в таком смысле действительно встречаются.
Информация и упорядоченность связаны, а поскольку все наши наблюдения и наше понимание энтропии говорят нам, что в природном, спонтанном, неуправляемом и незапрограмированном процессе упорядоченность будет уменьшаться, то тоже самое характеризует и информацию. Физик и инженер связи пожалуй не удивятся узнав что биологические процессы не сопровождаются увеличением полезной или функциональной (телеономической) информации и сложности. На самом деле, конечный результат любого биологического процесса, сопровождающегося передачей информации (т.е. всей наследственной изменчивости) есть сохранение или потеря этой генетической информации.
А это указывает на то, что первоначально информации была создана сверхестественным путем. Такой взгляд находится в полном согласии с библейской концепцией сбалансированного и цельного мира, сотворенного «очень хорошо», и распад которого начался только после грехопадения. Вот почему неизбежно существуют пределы изменчивости и почему креационист не должен беспокоиться о том, как много новых «видов» возникнет в будущем — однажды был положен предел количеству имеющейся функционально эффективной информации, и поэтому такой естественный процесс как мутирование не может увеличить ее первоначальный объем.
Заметим, что поскольку организмы были созданы для хотя бы одной миграции из центральной точки, и должны были заполнить пустые экологические ниши и преодолевать проблемы распадающейся и изменяющейся среды, было необходимым присутствие значительного потенциала изменчивости. Без такой заложенной в них генетической пластичности, большинство популяций не дожили бы до сегодняшнего дня. Следовательно, концепция биологического изменения, в каком-то смысле, предсказана библейской моделью, а не навязана ей только потому, что такое изменение произошло.
СОТВОРЕННЫЙ РОД
В писании подразумевается, что эта изначально сотворенная информация не появилась в форме одного «супервида», от которого отделялись все сегодняшние популяции в процессе постепенного «откалывания», но была сотворена в форме нескольких отдельных генофондов. Каждая группа организмов, размножающихся половым путем состояла, по-крайней мере, из двух особей. Поэтому
- Каждая первоначальная группа начинала свое существование с уже встроенной генетической информацией, которая явилась сырым материалом для практически всей последующей полезной изменчивости.
- Каждая первоначальная группа, вероятно, была генетически и репродуктивно изолированной от других подобных групп, но особи внутри своей группы могли свободно скрещиваться между собой. Поэтому, первоначальные сотворенные роды могли бы вполне подходить под современное определение «вида».4 На упомянутом примере собак, мы видели что такие «виды» могли раскалываться на две или более отчетливые подгруппы, которые затем могли расходиться (без добавления нового) и, в конце концов, могли сами приобретать характеристики «видов» — то есть репродуктивно изолироваться друг от друга, при этом свободно скрещиваясь внутри своей группы. Чем больше была изменчивость в первоначальном генофонде, тем легче могли возникать такие новые группы. Однако, каждый такой «раскол» уменьшал потенциал для дальнейшего изменения т.е. и тут присутствовал предел. Все потомки такого первоначального рода, который когда-то был видом, могли быть в конечном счете классифицированы, например, как одно большое семейство – более крупную таксономическую единицу.
Возьмем к примеру гипотетический сотворенный род А – настоящий биологический «вид», с огромным генетическим потенциалом. См. Рис. 1 (С целью упрощения, диаграмма не учитывает наличие двух особей каждого рода на борту ковчега – однако, главная идея при этом остается без изменений). Заметим, что А, как группа, даже может оставаться без изменений, равно как и каждая из подгрупп. Уход дочерних популяций в результате отщепления не обязательно означает вымирание родительской популяции. В случае человека, первоначальная группа не дивергировала настолько чтобы образовался новый вид.
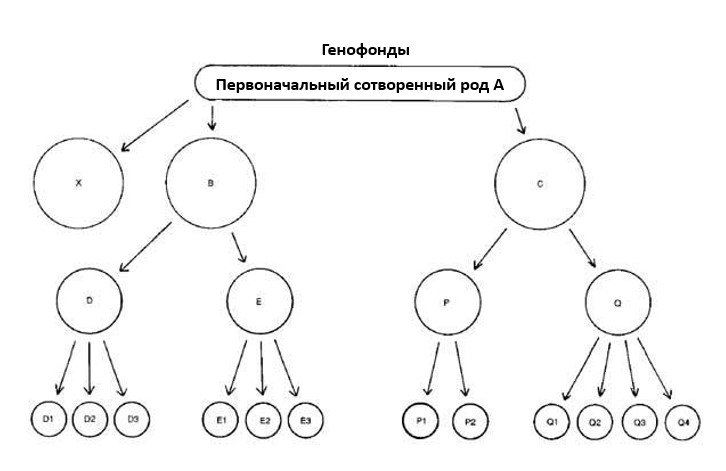
Рисунок 1. Разветвление дочерних популяций от первоначального сотворенного рода.
Отсюда получается, что D1, D2, D3, Е1, Е2, Е3, P1, P2, Q1, Q2, Q3 и Q4 все являются разными видами, изолированными репродуктивно. Однако, вся функционально- эффективная генетическая информация, содержащаяся в них уже присутствовала в А. (Предположительно, они также несли в себе мутационные дефекты).
Давайте предположим, что первоначальный сотворенный род А вымер, а вместе с ним и популяции X, B, C, D, Е, P и Q. (Но не D1, D2, и т. д.). Если Х нес часть первоначальной информации А, которая не представлена в B и C, тогда эта информация потеряна навсегда. Отсуда, несмотря на факт наличия многих «новых видов», которых первоначально не было, мы были свидетелями сохранения большей части информации, потерю некоторой информации и отсутствия новых добавлений помимо мутаций (вредных дефектов или просто бессмысленного «шума» в генетической информации). Все это – тот тип изменения информации, который не годится для демонстрации эволюции от простейших до человека.
Классификации на уровнях выше видового — это более или менее произвольные группировки, созданные для удобства и опирающиеся, главным образом, на сходствах или различиях в структуре. Вполне возможно, что сегодня D1, D2 и D3 могли бы быть классифицированы как виды, принадлежащие к одному роду, а Е1, Е2, Е3 как, например, виды другого рода. Также могло быть, что группы В и С могли различаться до такой степени, что их потомки оказались бы сегодня в разных семействах. Мы начинаем видеть некоторые проблемы для креациониста, который пытается разделить сегодняшних представителей сотворенных родов.
Живые существа могли бы быть отнесены к одному семейству, например, на основе сходства общего плана строения, в то время как они принадлежали бы к двум совершенно разным сотворенным родам. Это звучит как нота предостережения против использования морфологии как единственного критерия классификации, а также указывает на потенциальную ошибочность заявлений типа: «в данном случае барамин есть семейство; а в этом случае – таксономический род, и т.д.» (В креационизме, барамин – это общепринятый термин для «сотворенного рода».)
Простого решения для проблемы определения каждого из этих генетических взаимоотношений пока не существует- фактически, мы, наверное, никогда не сможем с определенностью узнать о них все. Скрещивание, эксперименты по оплодотворению in vitro и т.д. могут указывать на принадлежность к одному барамину, но отсутствие подобной генетической совместимости не доказывает что две группы не принадлежат к одному и тому же сотворенному роду. (См. вышеупомянутую дискуссию – генетические барьеры могут возникать в результате мутационных нарушений.) Однако, новейшие открытия дают нам возможность делать прямые сравнения между видами посредством анализа последовательностей ДНК, что открывает совершенно новый горизонт исследований. (Хотя, вопрос возможности широкого финансирования таких исследований в обществе, где доминируют эволюционные взгляды, остается открытым.)
Итак, что же нам сегодня сказать эволюционисту, который по понятным причинам, будет настаивать дать определение «сотворенному роду» или спросит как этот род опознать? Я выдвигаю на рассмотрение следующее:
«Группы живых организмов принадлежат к одному сотворенному роду, если они произошли из одного предкового генофонда.»
При таком определении нет необходимости говорить об «устойчивости сотворенных родов» по отношению к разновидностям сегодняшнего дня – по определению, новые роды не могут появиться.
Несмотря на то, что это определение является простым и очевидным оно аксиоматично. Оно бесстыдно круговое как свернувшийся броненосец и такое же непроницаемое, переводящее внимание на реальный вопрос генетического изменения. Вопрос не состоит в том, что такое барамин- вид, семейство или таксономический род? Вопрос в том, какие из сегодняшних популяций являются родственниками друг другу благодаря этой форме общего происхождения и происходят от того же сотворенного рода. Заметьте, что это повсеместно удалено из эволюционного понятия общего происхождения. По мере того как креационист движется взглядом в прошлое, двигаясь к истокам происхождения, он видит расширение генофонда. Когда эволюционист проделывает то же самое, он видит сжатие.
Как это бывает со всеми таксономическими вопросами, здравый смысл, наверное, снова будет играть главенствующую роль. Священное писание, данные по ископаемым окаменелостям, и здравый смысл, взятые вместе, не дадут креационистам «валить все в кучу» по мере движения назад во времени. Например, разумно считать (хотя и не всегда), что крокодилы и аллигаторы произошли из одного предкового генофонда, имевшего все функционально-эффективные гены, но вряд ли крокодилы, аллигаторы и страусы имели общий предковый генофонд, который содержал гены всех этих трех групп.
Ссылки и примечания
1. Lewin, R., Evolutionary theory under fire, Science 210(4472):883–887, 1980.
2. Lewin, ref. 1, p. 884.
3. Транспозоны или «прыгающие гены» , которые переносят генетическую информацию с одного участка ДНК на другой (в одном и том же организме, а иногда и в разных, как например в случае вирусов) в этой дискуссии не рассматриваются, поскольку работы в этой сфере «новой надежды» для эволюции не достигли достаточно зрелой фазы. Однако, необходимо заметить,что эти процессы, повсему видно, как и мутации являются случайными и ненаправленными процессамии, включающими перенос уже существующей информации.
4. Конечно, первоначальный «сотворенный род» мог быть представлен двумя или более отдельными популяциями способными к скрещиванию, но географически изолированными. Таким образом, факт их репродуктивной «изоляции» мог представить их , благодаря ультра-строгому использованию определения, одним видом «сотворенного» рода. Некоторые креационисты назвали бы этих изолятов «морфотипами», первоначально сотворенными внутри каждого барамина.
Если вам понравилась статья, поделитесь ею со своими друзьями в соц. сетях!
ВАМ БУДУТ ИНТЕРЕСНЫ ЭТИ СТАТЬИ:







