
Эволюция лошади
Автор: Матс Молен (англ. Mats Molén)
Источник: creation.com
Перевод: Алена Недоступ
Редактура: Елена Бондаренко, Владимир Силенок
Филогенетический ряд лошадей уже давно стал витриной эволюции. Но на самом деле этот ряд — лучший аргумент из палеонтологической летописи, который может быть представлен против эволюции.[1] Креационисты имеют различные мнения о том, состоит ли эта группа на самом деле из различных созданных родов. В этой статье рассматриваются некоторые из текущих проблем, и делается вывод, что группа, вероятно, включает три различных созданных рода, не включая всех животных, под названием гиракотерии. Сами гиракотерии, по-видимому, содержат несколько различных созданных родов животных, похожих на тапиров.
Окаменелости лошадей были найдены в осадочных слоях в начале третичного периода в отложениях эпохи, называемой эоценом (приблизительно 50 миллионов лет назад, согласно униформистской датировке). Их обычно называют[2] Eohippus или Hyracotherium (см. рис. 1).
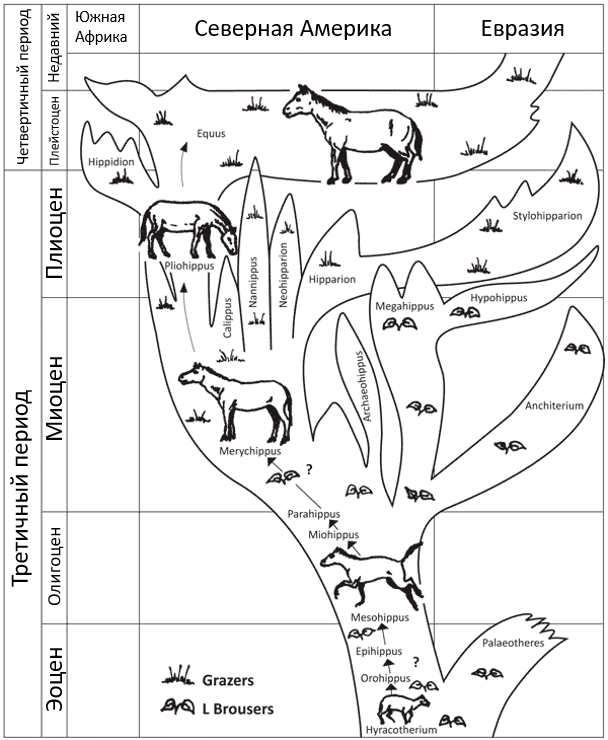
Рисунок 1. Эволюционное древо лошади, созданное Джорджем Гейлордом Симпсоном (George Gaylord Simpson) в 1951 году. Древо было позже упрощено,[5] но в последнее время стало еще более ветвистым и запутанным с добавлением большего числа представителей в результате открытия новых ископаемых находок (см. ссылку 2). Возможные эволюционные пробелы здесь отмечены знаком вопроса. Equus = современная лошадь.
Согласно теории эволюции, можно проследить эволюцию лошади на протяжении миллионов лет: как лошадь медленно становилась больше и сильнее (рис.1), теряла многие из своих пальцев (рис. 2), и как изменилась структура ее зубов, когда она перешла от рациона широколиственных растений, кустарников и деревьев к поеданию жесткой, сухой травы (пастьба) (рис. 3).[3],[4] Считается, что эволюция лошадей была обусловлена холодным и сухим климатом. Ранние лошади предположительно жили во влажных лесах, богатых растениями с пышной листвой. Их пальцы ног, четыре спереди и три сзади, вытянулись под разными углами, что помогало им не утонуть в болотистой почве. По мере того как климат становился суше, исчезали лиственные растения и образовывались огромные поля травы. Это заставило травоядных лучше бегать, чтобы спасаться от хищников.
Все лошади настолько похожи друг на друга, что были отнесены к одному семейству — Equidae. Из-за этого близкого сходства часто бывает трудно обнаружить какие-либо различия при изучении только ископаемых скелетов. Нужно проявлять осторожность при идентификации ископаемых позвоночных ещё и в том, что вариации в строении даже в пределах одного рода живых животных часто могут быть настолько велики, что они перекрываются с вариациями в других группах; например, существует много аналогий в строении зубов между различными плотоядными, даже если животные не относятся к одному роду (или иногда даже к одному семейству). Наиболее важные диагностические отличия между различными группами животных часто заключаются в строении мягких частей тела. Кроме того, многие находки ископаемых лошадей состоят только из зубов или частей челюстей.
Группы лошадей
В группе лошадей можно различить некоторых животных, которые могли бы представлять созданные роды, даже если у нас есть доступ только к ископаемым скелетам. Следующие факты, по-видимому, подтверждают такую интерпретацию.
В ряду лошадей есть как минимум два эволюционных пробела
а) первый пробел имеет место в таксоне Epihippus[8]
Окаменелости этого животного редкие и они напоминают окаменелости более ранних Орогипусов, Eohippus и других ранее идентифицированных видов гиракотерид.[9]
b) второй пробел имеет место в группе Parahippus[10] или сразу после нее.
Считается, что ранние виды парагипусов похожи на Miohippus и Mesohippus, а последний — на Merychippus; это лишь частично подтверждается находками ископаемых.[11] Кроме того, ископаемый материал для парагипусов является неполным.[12] Вероятно, можно было бы отнести различные части парагипусов к двум различным животным — Miohippus (рис. 4) и Merychippus.[13] Этот последний результат может быть также выведен из работы Cavanaugh et al.[14] потому что парагипус выказывает сходства с 14 из 18 видов лошадей. Таким образом, ступень «парагипус» в группе лошадей представляется смешанной группой неродственных ископаемых.
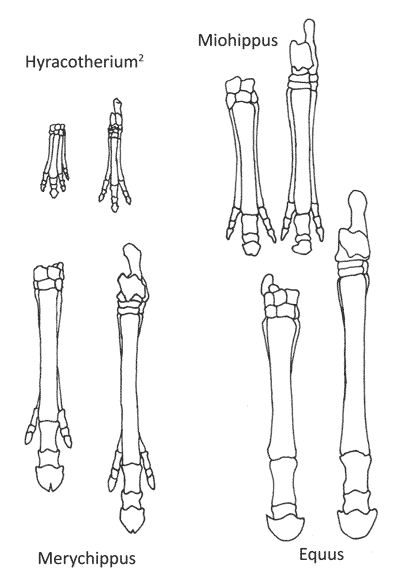
Рисунок 2. Ноги лошадей, которые воспринимаются как опора для эволюции. Левая нога в каждой паре на картинке — передняя, а правая — задняя.[6]
С 1989 года оспаривается монофилия таксона гиракотерий[9]
В 1992 году род гирактотерий был пересмотрен и отнесен к пяти животным, принадлежащим к разным семействам, из которых только одна группа считалась имеющей какое-либо отношение к лошадям.[15] Более поздние исследования перераспределили этих животных на десять различных родов и, по крайней мере, три семейства, из которых многие, как предполагается, не имеют ничего общего с рядом лошадей, но похожи, например, на тапиров (семейство Tapiromorpha).[9] Из рода гиракотерий один вид (angustidens) был переименован в Eohippus, а всем другим видам гиракторий, кроме одного, были даны новые родовые названия. Единственное животное, все еще сохранившее название гиракотерии (leporinum), больше не принадлежит к ряду лошадей, но считается принадлежащим к семейству палеотериевые, которые напоминают тапиров и носорогов.
«Ранние» лошади сохранились в слоях того же эволюционного возраста, что и несколько «более поздних» лошадей
Гиракторий/Eohippus и орогиппусы, например, появляются в палеонтологической летописи в одно время с Epihippus. Mesohippus и Miohippus появляются вместе с Merychippus и парагиппусами. Почти все другие лошади (за возможным исключением одной или двух) — парпгиппус, Merychippus, Pliohippus, Equus и, возможно, также Miohippus — находят в одно и то же время в окаменелостях.[16] (Но особенно в новых эволюционных схемах, различные имена были даны очень схожим животным, что даёт видимость эволюции, а также обеспечивает славу их первооткрывателям; см. примеры в Froehlich 2002[9] и MacFadden 2005[4]). Окаменелости гиракотерий (sic) также были найдены очень высоко в пластах (плиоцене), но эти находки были отвергнуты как переотложенные (т. е. эродированные и отложенные в более поздних слоях), несмотря на то, что геологические наблюдения не показывают никаких признаков того, что они были потревожены.[17] Таким образом, тот факт, что большинство лошадей жили почти в одно и то же время, подрывает их предполагаемую эволюцию.
«Переходные» формы между лошадьми с зубами, предназначенными для поедания листьев (Parahippus), и лошадьми с зубами, предназначенными для поедания травы (Merychippus), встречаются редко.[13]
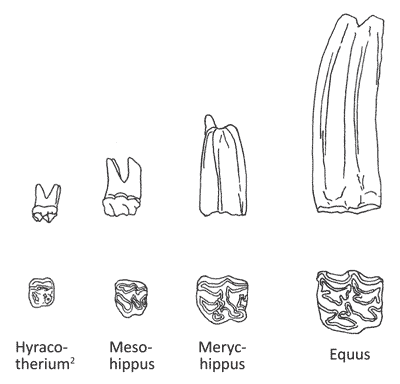
Рисунок 3. Строение зубов у листоядных (два слева) и травоядных лошадей (два справа).[7]
Зубы у поедающих листья лошадей имеют закрытые, очень узкие корни с небольшими отверстиями для их кровоснабжения и нервов; то есть это зубы, которые изнашиваются по мере взросления животного. Зубы у пасущихся (травоядных) лошадей имеют открытый корень с множеством кровеносных сосудов, которые снабжают зубы большим количеством питательных веществ, чтобы они могли расти в течение всей жизни животного; это называется гипсодонтия, что означает «зубы с высокой коронкой». Это изменение структуры зуба от бунодонта (зубы с низкой коронкой и закругленными бугорками) к гипсодонту (с высокой коронкой) — не является просто предполагаемой «микроэволюцией», а полным изменением конструкции, хотя это может и не показаться чем-то совершенно новым для тех, кто не знаком с конструкцией зуба.[18] Нет никаких доказательств того, что одна структура зуба меняется на другую, хотя это и предполагалось некоторыми авторами.[19] Некоторые животные ели и траву, и листья, но это не помогает объяснить превращение одного вида зубов в другой.
Три совершенно разных животных
Таким образом, животные, которые были истолкованы как различные лошади, легко идентифицируются как принадлежащие к трем совершенно различным родам животных, а не к различным промежуточным звеньям лошадей, которые предположительно произошли от одного и того же первоначального предка. Созданные роды, не считая всех членов гиракотериев, которые были переведены в новые семейства, должны более или менее соответствовать следующим трем группам (обратите внимание, что не все вновь названные лошади и не все члены боковых групп упомянуты ниже):
1. Eohippus (и многие ископаемые, которые ранее называлис гиракотериями, но классифицированые как принадлежащие к семейству Equidae с новыми родовыми названиями[9]), орогиппусы и Epihippus.
2. Mesohippus, Miohippus, некоторые парагиппусы и, вероятно, большинство лошадей, ответвляющихся от этих трех групп. (Ряд лошадей был перестроен и в него было добавлено много новых родов; например, клады Neohipparion, Nannippus и гиппарионы были перемещены ближе к парагиппусам и дальше от Merychippus,[4] в отличие от расположения на рисунке 1, поэтому мы не можем быть уверены, что классификация/группировка всех ископаемых правильна. Но лошади, ответвляющиеся от Merychippus на рис. 1, все еще классифицируются в подсемействе Equinae и поэтому объединяются в третью группу ниже. Но все эти детали не могут быть рассмотрены в данной статье).
3. Merychippus и те лошади, которые ответвились от этой группы, включая Pliohippus и всех более поздних лошадей (включая ветвь гиппарионов). (Обратите внимание, что в последних редакциях эволюции лошади есть два различных рода с именем Merychippus: I и II. Merychippus, следовательно, считается полифилетическим, то есть считается, что он эволюционировал дважды. Эти два рода были поставлены на разные эволюционные линии. Род I находится в исходном месте, ведущем к Equus, как видно из большинства диаграмм эволюции лошадей. Род II был отодвинут от линии, ведущей к Equus — он современен с парагиппусом в течение большей части своего существования — и считается предком ветви гиппариона, согласно MacFadden 2005.[4])

Рисунок 4. Две «лошади», Neohipparion (справа) и Miohippus (слева) из Музея естественной истории в Лос-Анджелесе.
Животные из 3-й группы относятся к одному подсемейству — Equinae.[20] Хотя, Cavanaugh et al.[10] обнаружив, что ископаемые животные могут быть разделены на подсемейства, проигнорировали это открытие и вместо этого построили свое собственное дерево эволюции лошади. Было бы нетрудно создать подобное дерево, просто расположив любое количество неродственных живых животных в ряд от малого до большого (рис.5).
Не существует эволюции лошади
Гипотеза Кавано (Cavanaugh), Вуда (Wood) и Уайза (Wise) о том, что ряд лошадей (включая род гиракотерий) показывает реальную (послепотопную) «микроэволюцию» (или линейное/прогрессивное видоизменение) на основании вышеуказанных результатов, несостоятельна, поскольку в эволюции лошадей нет прогрессии (за исключением, возможно, локальной), а данные показывают смесь различных животных, похожих на лошадей. Более того, статья Cavanaugh et al.[14] была основана главным образом на статистических данных из одного источника 1989 года (и некоторых дискуссиях из более поздних креационистских журналов), и она не квалифицировала различные находки гиракотерия. Кроме того, в феврале 2002 года была опубликована статья Фройлиха (Froehlich),[9] в которой были реклассифицированы все виды гиракотрий, примерно за год до срока публикации Cavanaugh 2003 et al. ICC.[14] Эта неясность в отношении находок гиракотерия также не была рассмотрена в статье Вуда (Wood ,2008 г.),[21] хотя Вуд ссылался на книгу Макфаддена (MacFadden) 1992 года,[22] который утверждал, что гиракотерий был не одним животным, а несколькими родами, принадлежащими к разным семействам. Уитмор и Уайз (Whitmore and Wise, 2008) даже использовали гиракотерий для установления ранней послепотопной датировки, и это животное, не являющееся лошадью, упоминается как первый член в ряду лошадей.[23]
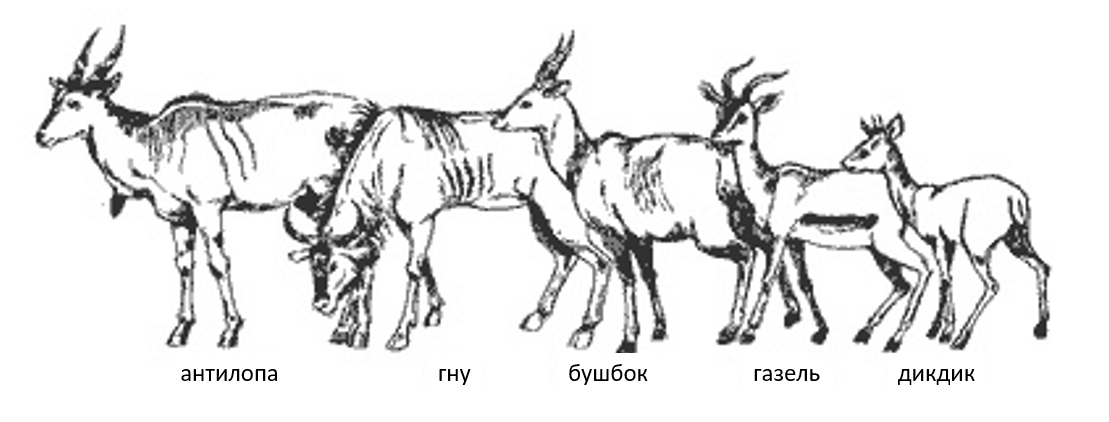
Рисунок 5. Слева направо — лось, гну, бушбок, газель и дикдик. Даже живущие сегодня животные могут быть организованы в гипотетический эволюционный ряд, поскольку вариации скелета в пределах одной группы животных часто перекрываются с вариациями в других группах в пределах того же семейства. Однако это не доказывает, что какое-либо отдельное животное эволюционировало в другое.
Фройлих,[9] который полностью переименовал большинство видов гиракотерий и поместил их в разные роды и семейства, использовал статистику, но также критиковал то, как статистика может быть неправильно использована в этом случае. Но, во любом случае, нельзя использовать статистику дизайна или использовать ограниченное количество данных (которые в этих случаях являются в основном зубами и челюстями), чтобы выяснить, как предположительно произошла эволюция, как это сделали вышеупомянутые авторы.[9],[14] Статистический анализ в этом случае не принимает во внимание функцию или завершенные/спроектированные живые существа, а может только сравнивать небольшие различия (см. также более критические точки зрения у Фройлиха). В этом случае большая часть статистического анализа была проведена на небольших различиях зубной эмали/структуре и челюстях, и очень мало работы было сделано на других частях тела. Это искажает интерпретацию данных таким же образом, как если бы, например, мы собрали статистическое разнообразие по 75 параметрам внешнего вида глаз осьминогов и людей — анализ, вероятно, показал бы, что мы произошли от осьминогов.
Хотя легко обсуждать и критиковать отдельные находки или одно место, где были найдены окаменелости, согласно всем имеющимся данным, существует три группы животных, которые близко соответствуют подсемейству Equidae, и только подсемейство Equinae, по-видимому, представляет лошадей. Дискуссия о критериях границ Потопа и послепотопного периода, основанная на эволюции лошади, например, Cavanaugh et al. 2003[14] и Wood 2008,[21] следовательно, должны основываться на иных критериях, чем предполагаемая послепотопная «микроэволюция» лошади в результате изменения окружающей среды, как это предлагается общей эволюционной историей (см. другие критерии границ потопа в Oard 2007[24]). Не было также никакой реальной среды, где эти животные могли бы жить, кроме обширных пустынь — большинство окаменелостей найдено в осадочных отложениях, которые свидетельствуют о Потопе, но нет никаких свидетельств растительного покрова, который мог бы прокормить большие стада животных, и нет надлежащей почвы.25 Также нет доказательств изменений в окружающей среде, на чем настаивают эволюционисты и Cavanaugh et al.[14] и Wood,[21] основываясь на умозрительных интерпретациях.
В случае с лошадью, размер тела определял, как быстро животные утонули, были перенесены и захоронены, а затем иногда подвергались действию эрозии и повторному депонированию во время Потопа или сразу после него. Это должно было произойти до того, как континентальная среда снова стала пригодной для жизни и живые животные вновь заселили ее. Мелкие животные с похожим строением тела обычно распадаются и тонут быстрее, чем крупные животные, и более мелкие кости также легче переносятся течениями после того, как опустились на дно. Кроме того, во время послепотопных катастроф живые животные могли быть захоронены вместе с переотложенными, мертвыми, не окаменевшими или частично окаменевшими останками животных, захороненными во время Потопа.
Вывод
Изучение ископаемых лошадей показывает, что существует по крайней мере три группы животных в пределах семейства лошадиных Equidae, в дополнение к некоторым неродственным животным, таким как тапиры. Три группы Equidae близко соответствуют различным подсемействам Equidae и могут рассматриваться как три отдельных созданных рода. Большинство из этих различных родов жили (или фактически были похоронены!) почти в то же самое время и не выказывают больших прогрессивных изменений, по крайней мере в эволюции лошади, кроме простого общего увеличения размера.
Никто не объяснил, как предположительно могли развиться новые, специализированные типы зубов, и это, скорее, случай разумного замысла, а не «микроэволюции» (вариации внутри вида, как предполагают различные креационисты) или «макроэволюции» (новые виды организмов, как предполагают эволюционисты).
Гипотеза Cavanaugh et al. (2003)[14] об интрабараминальной изменчивости всех животных, принадлежащих к Equidae (или животных, которых они поместили в Equidae, даже если эволюционисты поместили некоторых из них в разные семейства), не вполне подтверждается имеющимися доказательствами и поэтому должна быть оставлена.
Дополнение
По мнению Джулиана Хаксли (Julian Huxley), возможно, одного из самых выдающихся эволюционистов прошлого века, для эволюции современной лошади требовалось не менее миллиона положительных мутаций. Он полагал, что, в общей сложности, на 1000 мутаций происходит максимум одна положительная. С помощью этих величин Хаксли рассчитал вероятность того, что лошадь эволюционировала из одного единственного одноклеточного организма составляет 1 к 103 000 000. Однако он верил, что естественный отбор сможет решить эту проблему.[26] Но эта вера не помогла ему в конце концов, и не поможет ни одному другому эволюционисту, так как этот расчет основан на происхождении положительных мутаций еще до того, как естественный отбор начал бы работать. Если бы все электроны во Вселенной (около 1080) участвовали в 1012 реакциях каждую секунду, в течение 30 миллиардов лет – предел, который определяется эволюционистами, как максимальный для возраст вселенной, то все еще не было бы больше 10110 возможных взаимодействий, что все еще очень далеко от расчетов Хаксли.[1]
Ссылки и примечания
1. Эта статья отчасти основана на английском переводе, сделанном Руди Арбеллом, статьи Molén, M., Vårt ursprung? (Our Origin), XP-media, Haninge, 2000.
2. Название гиракотерий больше не используется ни для одного представителя семейства Equidae, но ископаемым этого рода, которые все еще классифицируются в Equidae, были даны семь новых родовых названий: Sifrhippus, Minippus, Arenahippus, Xenicohippus, Eohippus, Plilolophus и Protorohippus.
3. Некоторые лошади с зубами, предназначенными для поедания травы, вероятно, в основном ели листья и другие мягкие части растений. См.: Morell, V., Dietary data straight from the horses mouth, Science 283:773, 1999; and MacFadden, B.J. et al., Ancient Diets, Ecology, and Extinction of 5-Million-Year-Old Horses from Florida, Science 283:824–827, 1999.
4. MacFadden, B.J., Fossil horses—evidence for evolution, Science 307:824–827, 2005.
5. Simpson, G.G., Horses, Oxford University Press, New York 1951.
6. The picture is based on drawings from: Steinmann, G. and Döderlein, L., Elemente der Paläontologie (Elements of Paleontology), Wilhelm Eng-elmann Publishing, Leipzig, 1890; Osborn, ref. 11; and Scott, ref. 12.
7. The picture is based on drawings from: Steinmann and Döderlein, ref. 6; Osborn, ref. 11; Mutvei, H. and Stuenes, S., Paleontologi: Kompendium för grundkurs i paleontologi (Paleontology: Compendium for Basics in Paleontology), Uppsala University, Institute of Paleontology, Uppsala, 1983; and Granger, W., A revision of American Eocene horses, Bulletin of the American Museum of Natural History 24:221–264, 1908.
8. MacFadden, B.J., Cladistic analysis of primitive equids, with notes on other perissodactyls, Systematic Zoology 25:1–14, March 1976; and Simpson, G.G., Horses, Oxford University Press, New York, 122–123, 203, 1951.
9. Froehlich, D.J., Quo vadis eohippus? The systematics and taxonomy of the early Eocene equids (Perissodactyla), Zoological Journal of the Linnean Society 134:141–256, February 2002.
10. Abel, O., Paläobiologie und Stammesgeschichte (Paleobiology and the History of Phylogeny), Gustav Fisher Publishing, Jena, pp. 284–290, 1929; and MacFadden, B.J. et al., Sr-isotopic, paleomagnetic, and biostratigraphic calibration of horse evolution: evidence from the Miocene of Florida, Geology 19:242–245, 1991.
11. Osborn, H.F., Equidae of the Oligocene, Miocene, and Pliocene of North America: iconographic type revision, Memoirs of the American Museum of Natural History 2:74–75, 1918. Например, одна из самых «примитивных лошадей» — Parahippus cognatus — предположительно эволюционировала как один из последних видов, в то время как одна из наименее примитивных — P. Coloradensis — эволюционировала одновременно с первым видом Parahippus.
12. Scott, W.B., A History of Land Mammals in the Western Hemisphere, 2nd ed., Macmillan Publ. Co., NY, p. 409, 1937.
13. В группе Parahippus есть некоторые находки, которые, как полагают некоторые исследователи, являются промежуточными формами между Parahippus и Merychippus (например, ссылаясь на изображения в Osborn, ссылка 11, где некоторые из наименее «развитых» «лошадей» возникли среди последних «лошадей»; см. стр. 74-75). Эти находки состоят только из зубов и частей челюстей, которые трудно привязать к определенному скелету. Следовательно, эти зубы и челюсти можно было бы разделить на Parahippus и Merychippus, а не на какие-либо промежуточные формы между этими двумя животными.
14. Cavanaugh, D.P., Wood, T. and Wise, K.P., Fossil equidae: a monobaraminic, stratomorphic series; in: Walsh, R.E. (Ed.), Proceedings of the Fifth International Conference on Creationism, Creation Science Fellowship, Pittsburgh, PA, p. 143–149, 2003.
15. MacFadden, B.J., Fossil Horses, Cambridge University Press, Cambridge, pp. 92–103, 1992. See also Scheven, J., Mega-Sukzessionen und Klimax im Tertiär: Katastrophen zwischen Sintflut und Eiszeit (Megasuccessions and Climax in the Tertiary: Catastrophes Between the Flood and the Ice Age.), Wort und Wissen, Neuhausen, 1988.
16. Scott, ref. 12, p. 408 and Osborn, ref. 11, p. 74; Barnhart, W.R. A Critical Evaluation of the Phylogeny of the Horse, ICR, 1987; MacFadden, ref. 15, pp. 255–257; Storer, J.E. and Bryant, H.N., Biostratigraphy of the Cypress Hills Formation (Eocene to Miocene), Saskatchewan, Journal of Paleontology 67:660–669, 1993; Gould, S.J. and Eldredge, N., Punctuated equilibrium comes of age, Nature 366:223–227, 1993; and Scheven, ref. 15.
17. Barnhart, ref. 16, pp. 140–143.
18. Personal communication with Dr Jack Cuozzo.
19. Sarfati, J., The non-evolution of the horse, Creation 21(3):28–31, 1999.
20. Gromova, V.I., Mammals in: Fundamentals of Paleontology: A Manual for Paleontologists and Geologists of the USSR, vol. 13, Orlov, Y.A. (Ed.), Jerusalem, English translation, Israel Program for Scientific Translations, p. 396, 1968.
21. Wood, T., Horse fossils and the nature of science, 2008; answersingenesis.org/articles/am/v3/n4/horse-fossils.
22. MacFadden, ref. 15, pp. 92–103.
23. Whitmore, J.H. and Wise, K.P., Rapid and early post-Flood mammalian diversification evidenced in the Green River Formation; in: Walsh, R.E. (Ed.), Proceedings of the Sixth International Conference on Creationism, Creation Science Fellowship, Pittsburgh, PA, pp. 449–457, 2008.
24. Oard, M.J., Defining the Flood/post-Flood boundary in sedimentary rocks, Journal of Creation 21(1):98–110, 2007.
25. Molén, M., Mountain building and continental drift; in: Walsh, R.E. (Ed.), Proceedings of the Third International Conference on Creationism, Creation Science Fellowship, Pittsburgh, PA, pp. 353–367, 1994.
26. Huxley, J., Evolution in Action, Chatto and Windus, London, pp. 47–48, 1953.
Если вам понравилась статья, поделитесь ею со своими друзьями в соц. сетях!
ВАМ БУДУТ ИНТЕРЕСНЫ ЭТИ СТАТЬИ:







