
Дизайн возвратного гортанного нерва у млекопитающих – совсем не плохой дизайн
Автор: Др. Джерри Бергман (англ. Jerry Bergman)
Источник: creation.com
Перевод: Алена Недоступ
Редактура: Елена Бондаренко, Владимир Силенок
Распространенное утверждение эволюционистов состоит в том, что у млекопитающих возвратный гортанный нерв был плохо спроектирован потому что он проходит вниз минуя гортань, далее вокруг аорты и, наконец, обратно к гортани. Они считают, что гораздо более эффективным был бы намного более короткий путь непосредственно к гортани. Приведенный ниже анализ показывает, что причины более длинного маршрута включают в себя как ограничения развития, так и конструктивные ограничения. Кроме того, доказательства разумного замысла такой схемы очевидны и убедительны.
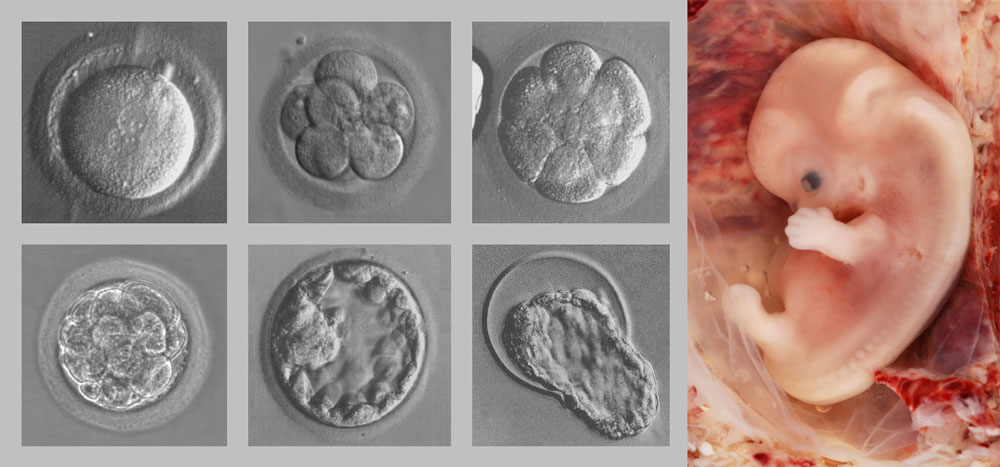
Рисунок 1. На самых ранних стадиях развития эмбриона необходимо удлинение, чтобы, в конечном итоге, изменить сферическую структуру в нормальное, вытянутое человеческое тело. На каждой стадии развития организм должен быть живым и также нуждается в продуманных модификациях по мере своего развития. Credit: Zygote: Jrockley and Embryo: Ed Uthman, MD: wikimedia.org
Эволюционисты обычно утверждают, что человеческое тело плохо спроектировано, и это доказывает, что оно не было разумно спроектировано, а скорее сколочено как попало неразумным процессом эволюции. Одним из наиболее распространенных примеров плохой конструкции, приводимых сегодня эволюционистами, является возвратный гортанный нерв (RLN, от англ. recurrent laryngeal nerve), который управляет мышцами гортани (голосового аппарата). Дарвинисты часто утверждают, что эволюция доказана потому, что примеры «плохого или, по крайней мере, очень загадочного дизайна могут накапливаться бесконечно», и одним из лучших примеров является
«…возвратный гортанный нерв, который соединяет мозг с гортанью и позволяет нам говорить. У млекопитающих этот нерв не идёт прямым путём от мозга к горлу, но, вместо этого, спускается в грудную клетку, делает петлю вокруг аорты около сердца, а затем возвращается в гортань. Это делает его в семь раз длиннее, чем нужно!»[1]
Главный аргумент заключается в том, что гортанный нерв плохо спроектирован, потому что он не проходит по кратчайшему пути к гортани, что также верно для многих других нервов. Примерами могут служить зрительные нервы, которые не проходят кратчайшим путем к затылочной доле мозга, а, вместо этого, пересекаются в хиазме (перекрёст зрительных нервов) по тем причинам, которые, как теперь известно, являются очень вескими, и связанными с оптимальном дизайном.[2]
Аналогичным образом, за исключением правой и левой височных ветвей лицевого нерва, которые управляются обеими сторонами мозга, нервы, отходящие от правого полушария мозга иннервируют левую сторону тела.
Профессор Ричард Докинз утверждает, что ВГН также является доказательством эволюции человека от рыбы. Он пишет, что во время предполагаемой эволюции млекопитающих
«…шея вытянулась (у рыб нет шеи), а жабры исчезли, некоторые из них превратились в полезные вещи, такие как щитовидная и паращитовидная железы, а также различные другие части, которые вместе образуют гортань. Эти полезные вещи, включая части гортани, получили свое кровоснабжение и нервные связи от эволюционных потомков кровеносных сосудов и нервов, которые давным-давно служили жабрам в упорядоченной последовательности».[3]
Однако не существует никаких научных доказательств того, что жабры «превращаются в полезные вещи, такие как щитовидная и паращитовидная железы». Жаберные клетки очень сильно отличаются от эндокринных клеток. Докинз делает вывод:
«По мере того как предки млекопитающих эволюционировали все дальше и дальше от своих предковых рыб, нервы и кровеносные сосуды оказывались натянутыми и растянутыми в загадочных направлениях, что нарушило их пространственные отношения друг ко другу. Грудь и шея позвоночных превратились в сплошное месиво, в отличие от аккуратной, симметричной, последовательной повторяемости рыбьих жабр. И возвратные гортанные нервы стали более чем обычно преувеличенными жертвами этого искажения».[3]
Любой, кто изучал анатомию человека, знает, что у позвоночных грудная клетка и шея — это не «месиво», а хорошо продуманная, сложная, функциональная система.
Анатомия ВГН
Как мы покажем далее, левый ВГН, который иннервирует гортань млекопитающих и внутренние мышцы надгортанника, имеет анатомическую траекторию, очень отличную от того, что можно было бы сперва ожидать и у этого есть веские причины. В отличии от утверждения Дональда Протеро (Prothero), ВГН не избегает «прямого пути между мозгом и горлом», а «вместо этого спускается в грудную клетку, петляет вокруг аорты около сердца, а затем возвращается в гортань».[1] Скорее блуждающий нерв проходит от шеи вниз к сердцу, а затем ВГН ответвляется от блуждающего нерва чуть ниже аорты (большая, главная артерия, идущая вверх от левого желудочка сердца и затем идущая вниз по брюшной полости). Наконец, как будет подробно описано ниже, эти ветви поднимаются вверх, чтобы иннервировать несколько органов, некоторые из которых находятся вблизи того места, где они ответвляются от блуждающего нерва.[4]
Нервные сигналы, управляющие различными функциями организма, идут от головного мозга вниз по позвоночнику или черепным нервам, а затем разветвляются, соединяясь с органами, которые они обслуживают. Гортань расположена в области шеи, а у эмбриона правый гортанный нерв ответвляется от блуждающего нерва в области шеи.[5]
Однако у взрослых людей, левый блуждающий нерв, вместо того, чтобы идти прямым путем от блуждающего нерва к гортани в области шеи, перемещается вниз по шее в грудную клетку рядом с сердцем, чтобы обеспечить холинергическую иннервацию многочисленных внутренних органов. Гортанный нерв ответвляется от блуждающего нерва в области грудной клетки, затем делает петлю под задней стороной аорты чуть выше сердца и, наконец, возвращается к гортани. По этой причине его называют левым возвратным гортанным нервом. Напротив, правый гортанный нерв петляет вокруг подключичной артерии, а затем поднимается вверх к гортани. Следует отметить тот факт, что более длинный левый ВГН прекрасно работает в гармонии с правым ВГН, что исключает утверждение о том, что это плохая или ошибочная конструкция.
Причины такого дизайна
Наиболее логичной причиной такого дизайна является то, что он обусловлен ограничениями в развитии. Эмбриолог, профессор Эрих Блехшмидт (Erich Blechschmidt) писал, что кажущаяся плохая конструкция возвратного гортанного нерва у взрослых обусловлена «необходимыми последствиями динамики развития и не должна интерпретироваться … как исторический перенос» эволюции.[6] Сконструированные человеком устройства, такие как радио и компьютеры, не должны функционировать до тех пор, пока их сборка не будет завершена. Напротив, живые организмы должны функционировать, причем хорошо функционировать, чтобы процветать на каждой стадии развития от зиготы до взрослого человека (рис.1). Эмбрион в целом также должен быть полностью функционирующей системой в своей специфической среде в течение каждой секунды своего развития. По этой причине анатомия взрослого человека может быть понята только в свете развития от зиготы к взрослому человеку:
«Путь для нервных волокон обычно определяется иннервируемыми органами и поэтому прокладывается извне. Мы должны предположить, что субмикроскопические движения материала (т. е. молекулярные) имеют решающее значение для этого процесса; а именно, что упорядоченные метаболические движения работают таким образом, который определяет форму зарождающегося образца иннервации».[7]
Аналогия, которую Блехшмидт использует, чтобы прояснить свой аргумент, — это течение реки, которое
«…не может быть объяснено на основе знания его источников, его притоков или конкретных мест расположения гаваней в его устье. Только общие топографические обстоятельства определяют течение реки».[7]
Именно из-за различий в топографическом ландшафте тела млекопитающих «ход нижнего гортанного нерва весьма неоднозначен», а незначительные анатомические различия являются обычным явлением.[8] Этот факт был задокументирован серией вскрытий шеи, выполненных на 90 человеческих трупах через 48 часов после смерти для изучения анатомии правого и левого возвратных гортанных нервов. Вскрытие показало, что путь этих нервов иногда отличался от того, что показано в стандартной медицинской литературе, иллюстрируя аналогию Блехшмидта.[9] Интересным результатом дизайна ВГН является то, что для врача он может быть индикатором сосудистого заболевания, т. е. увеличение аорты или подключичной артерии, вызванное аневризмой, может сжимать левый ВГН, вызывая дисфункцию голоса.[10] В результате такая дисфункция может свидетельствовать об аневризме.
Аспекты развития
Развитие человеческого тела начинается со сферы, называемой бластоцистой, и постепенно все более вытягивается по мере своего развития. Некоторые структуры, такие как сонный проток (проток, соединяющий у эмбриона сонные артерии и дуги аорты), просто уничтожаются во время развития, в то время как другие устраняются и позже заменяются по мере созревания плода. Другие структуры, включая возвратный гортанный нерв, перемещаются вниз по мере развития. Причина этого движения заключается в том, что формирование шеи и удлинение тела, которое происходит во время развития плода, заставляет сердце опускаться из шейного расположения вниз в грудную полость.[11]
В результате этого, различные артерии и другие структуры должны быть удлинены, поскольку органы перемещаются таким образом, чтобы оставаться функциональными на протяжении всей этой фазы развития. Правый возвратный гортанный нерв переносится радикально вниз, потому что он петлей проходит под IV дугой, которая развивается в правую подключичную артерию, и таким образом вынужден двигаться вниз вместе с ней.[12]
В тех случаях, когда правая IV дуга рассасывается, правый возвратный гортанный нерв не петляет, а соединяется непосредственно с гортанью.[9] Левый гортанный нерв петляет вокруг артериальной связки (VI дуга, небольшая связка, прикрепленная к верхней поверхности легочного ствола и нижней поверхности дуги аорты) с левой стороны дуги аорты и, таким образом, должен двигаться вниз по мере удлинения грудной полости. Блехшмидт отмечает, что:
«Не может существовать органа, который не функционировал бы в процессе своего развития. Эта аксиома применима и к нервной системе. Нервная система выполняет свои последующие функции за счет своих предыдущих функций роста».[13]
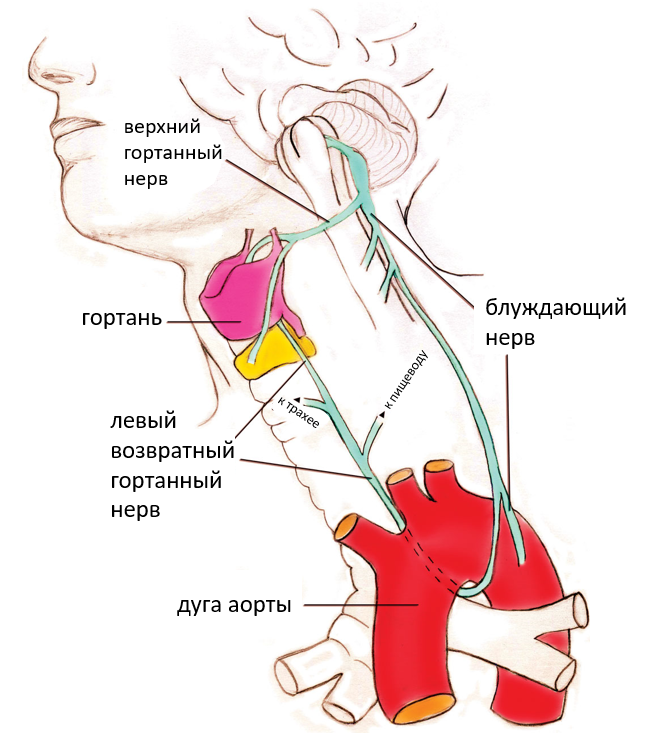
Рисунок 2. Схема левого возвратного гортанного нерва, который ответвляется от блуждающего нерва. Обратите внимание, что верхний гортанный нерв соединяется непосредственно с гортанью, а возвратный гортанный нерв иннервирует несколько структур, возвращаясь обратно к гортани (Illustration by Susan Windsor. Copyright © 2010 Institute for Creation Research. Все права защищены. Используется по разрешению)
В результате нисходящего движения «сердца и исчезновения некоторых аортальных дуг, ход возвратных гортанных нервов становится различным на правой и левой сторонах».[11] До этого сдвига эти два нерва обслуживают шестую висцеральную дугу, но по мере того, как сердце опускается, левый нерв огибает шестую аортальную дугу, а затем снова поднимается к гортани, что объясняет их возвратный ход. Эти нервы не могут быть ни уничтожены, ни заменены, потому что нерв должен функционировать на каждой стадии развития плода. Сэдлер (Sadler) объясняет, что на правой стороне
«…дистальная часть шестой аортальной дуги и пятая аортальная дуга исчезают, возвратный гортанный нерв движется вверх и прикрепляется к правой подключичной артерии. Слева нерв не движется вверх, так как дистальная часть шестой аортальной дуги сохраняется в виде артериальной связки».[14]
Это движение, по-видимому, предназначено для перетаскивания левого ВГН вниз по мере удлинения брюшной полости. Сэдлер также отмечает, что из-за того, что
«…мускулатура гортани берёт начало от мезенхимы четвертой и шестой висцеральных дуг, все мышцы гортани иннервируются ветвями десятого черепного нерва, т. е. блуждающего нерва. Верхний гортанный нерв иннервирует производные четвертой висцеральной дуги, а возвратный гортанный нерв — производные шестой висцеральной дуги».[15]
Тело должно работать как живая функциональная единица, требующая связок и внутренних соединений, чтобы скреплять между собой различные взаимосвязанные структуры, одновременно позволяя расти и перемещаться телу/органу, необходимого для гибкости требуемой для повседневной деятельности. Для гортанного нерва артериальная связка функционирует подобно подъязычной кости, позволяя двигаться. Нервы обычно не могут быть разорваны во время эмбрионального развития, а затем заново вырасти где-то еще. Тело так же не может разорвать нервы, чтобы позволить перемещению существующих нервов к другому месту, где они снова соеденятся (рис.2).
Существуют и другие случаи, когда один нерв рано отщепляется и обеспечивает прямую иннервацию, а другой идет, как кажется, окольным путем. Одним из примеров является диафрагмальный нерв (волокна с3, с4, с5), который отходит от области шеи и спускается к диафрагме. Это необходимая траектория, так как перикард и диафрагма возникают в поперечной перегородке (толстой массе черепной ткани, которая дает начало некоторым частям диафрагмы и вентральной брыжейке переднего отдела) в шейной области эмбриона на раннем этапе развития. Затем он мигрирует каудально (в направлении к копчику), поскольку эмбрион увеличивается за счет дифференцированного роста области головы и грудной клетки, увлекая с собой нерв. Диафрагма не могла бы развиваться поэтапно, так как частично сформированная диафрагма с дефектом приводит к несовершенному отделению грудной клетки от брюшной полости, и даже небольшой дефект приводит к образованию грыжи, т. е. протрузии органов брюшной полости в грудную клетку из-за дефекта в диафрагме – что либо сдавливает легкие, либо приводит к ущемлению кишечника.
По всем этим причинам приведенные ниже утверждения Протеро (Prothero) являются неверными и плохо осмысленными:
«Эта конструкция не только расточительна, но и … причудливый путь этого нерва замечательно объясняется с точки зрения эволюции. У рыб и ранних эмбрионов млекопитающих, предшественник возвратного гортанного нерва прикрепляется к шестой висцеральной дуге, глубоко в области шеи и тела».[1]
Сходство ВГН у всех позвоночных, включая рыб и млекопитающих, обусловлено сходным развитием эмбриона и плода в результате ограничений развития и сходной морфологией во взрослом состоянии. Это не связано с тем, что мы произошли от рыбы, как утверждает Протеро, что ни ископаемые ни другие свидетельства не подтверждают.
Другие функции ВГН
Кроме того, ветви возвратного гортанного нерва служат нескольким другим органам, включая верхнюю часть пищевода, трахею, нижнюю полость глотки и рожково-перстневидную связку, обеспечивая как моторные, так и сенсорные ветви, которые требуют данной существующей конструкции.[16],[17] Такое расположение позволяет структурам ниже гортани получать сигналы несколько раньше, чем гортань, чтобы подготовиться к активации гортани, когда такая функция неизбежна.
Кроме того, «от гортанной ветви отделяются и другие ветви, прежде чем прикрепиться к гортани на разных уровнях».[16] Нейроанатомы описывают иннервацию гортани как очень сложную, и они до сих пор пытаются определить специфические цели ее нервных ветвей. Тот факт, что от левого ВГН также отделяются некоторые волокна, которые соединяются с сердечным сплетением, тоже является весьма показательным для ограничений развития, поскольку нерв должен иннервировать как гортань (в шее), так и сердце (в груди).
Сложный вопрос, который до сих пор изучается, заключается в том, как невероятно сложная нервно–мышечная система, состоящая из нервных волокон и мышц гортани, возникает из нервного гребня и дорсальных сомитов соответственно у эмбриона на раннем этапе развития и затем мигрирует антериально в свое конечное местоположение. Утверждение о том, что ВГН плохо спроектирован, без объяснения того, как дизайн этой нервной структуры, ее функция, ее первичное происхождение и связи в мозге развивались от эмбриона до взрослого человека — просто бессмысленно. Таким образом, утверждение о том, что нерв должен без всякой причины удалиться от артериальной связки, является необоснованным.
Конструкция окольного пути
Некоторые иннервации гортани направляются непосредственно к ней, включая сенсорный внутренний гортанный нерв и двигательный внешний гортанный нерв. Два других нерва, левый и правый верхние гортанные нервы, ответвляются близко к гортани, чтобы обеспечить этой структуре прямую иннервацию. Верхний гортанный нерв ответвляется от блуждающего нерва в середине узловатого ганглия и получает ответвление от верхнего шейного ганглия симпатической нервной системы.[18]
Помимо причин развития, у обходного пути ВГН существуют и потенциальные преимущества перекрытия сенсорной и моторной иннерваций, когда один из нервов немного длиннее. Лучшее понимание иннервации гортани поможет нам понять необходимость немного более длинного маршрута для нерва, но сильный намек дает тот факт, что эти два нерва регулируют различные голосовые реакции.

Рисунок 3. Жираф иллюстрирует экстремальный пример необходимой длины левого возвратного гортанного нерва, однако система функционирует очень эффективно. Многие другие сложные конструктивные особенности также необходимы для создания знаменитой длинной шеи жирафа. (Photo by: Hollie Jeans, SXC)
Верхний гортанный нерв делится на внутреннюю и внешнюю ветви. Внешняя ветвь контролирует внутреннюю гортанную мышцу, перстнещитовидную мышцу[19] а также иннервирует мышцы, ответственные за увеличение высоты тона голоса. Различные другие ветви возвратного гортанного нерва иннервируют мышцы, ответственные за такие функции, как снижение высоты тона голоса, контроль громкости и утомляемость голоса. Три основные ветви ВГН иннервируют несколько мышечных пучков, включая щиточерпаловидную мышцу, заднюю перстнечерпаловидную мышцу и боковую перстнечерпаловидную мышцу. Повреждение нервов, иннервирующих эти мышцы, влияет на артикуляцию, и когда артикуляция нарушается, речь воспринимается как «невнятная» или «искаженная».[20]
Паралич верхнего гортанного нерва (нерва не идущего окружным путем) вызывает затруднения в повышении громкости голоса, производя высокий тон, и приводит к утомлению голоса и неспособности петь, потому что голосовые связки теряют свой нормальный тон и не могут достаточно удлиняться. Напротив, паралич одной или более из трех ветвей возвратного гортанного нерва может привести к слабому голосу, который звучит как у Микки-Мауса. В тяжелых случаях это может привести к параличу голосовых связок.[19]
Одному пациенту, который пострадал от травматического разрыва дуги аорты в автомобильной аварии, потребовался аортальный трансплантат, который оставил его с поврежденным левым ВГН. Единственным неблагоприятным результатом было то, что его голос был слаб, но его артикуляция оставалась неизменной. Он прекрасно говорит, но не может управлять своим голосом из-за того, что мышцы гортани обладают множественной иннервацией, а их совокупность, как единое целое, контролирует функцию голоса (интервью с доктором Видж Содера (Vij Sodera)).
Еще одна причина, по которой ветви гортанного нерва (обе из которых отходят от блуждающего нерва) расположены как выше, так и ниже гортани, заключается в том, что эта конструкция позволяет сохранить некоторые функции, если одна из них прерывается. Резервный путь также обеспечивает некоторую поддержку в случае повреждения одного из нервов.
Наконец, несколько исследований показали, что существующий путь занимает «относительно безопасное положение в борозде между трахеей и пищеводом»,[21] что делает его менее подверженным повреждениям или травмам, чем если бы у него был прямой путь.[22]
ВГН у жирафов
Жираф является любимым примером у тех, кто утверждает, что ВГН плохо спроектирован, (рис.3). Протеро писал, что ВГН жирафа «дважды пересекает всю шею, так что его длина составляет пятнадцать футов (из которых четырнадцать футов не нужны!)».[1] Докинз же утверждает, что у людей
«…путь, пройденный возвратным гортанным нервом, представляет собой крюк, возможно, в несколько дюймов. Но у жирафа это уже не шуточный крюк на много футов, возможно, около 15 футов у крупной взрослой особи!»[3]
Докинз добавляет, что «длина окольного пути, пройденного возвратным гортанным нервом», требует от команды анатомов одновременной работы на различных участках нерва, чтобы выделить ВГН, и это, как он отмечает, является «трудной задачей, которая, насколько нам известно, не выполнялась с тех пор, как Ричард Оуэн (Richard Owen), великий анатом викторианской эпохи, сделал это в 1837 году». Трудность заключается в том, что ВГН «очень узкий, даже нитевидный на участке, идущем кверху» и, следовательно,
«…его легко не заметить в запутанной паутине мембран и мышц, окружающих трахею. Во время своего нисходящего путешествия нерв (в этом месте он соединен с большим блуждающим нервом) проходит в нескольких дюймах от гортани, которая является его конечным пунктом назначения. И все же он проходит вниз по всей длине шеи, прежде чем развернуться и снова подняться вверх … я обнаружил, что мое уважение к Ричарду Оуэну (злейшему врагу Дарвина) растет. Креационист Оуэн, однако, не смог сделать очевидного вывода. Любой разумный конструктор по пути вниз обрубил бы гортанный нерв, заменив многометровый путь на путь в несколько сантиметров».[23]
Многие из описанных выше замечаний, касающихся людей, также применимы к жирафам. У эмбриона жирафа отсутствует шея, и он удлиняется, как и человеческий эмбрион, только гораздо больше, потому что его шея спроектирована так, чтобы быть намного длиннее, чем у человека. Следовательно, и ВГН перемещается вниз, когда шея удлиняется. Докинз утверждает, что шея жирафа удлинилась за время эволюции, хотя никаких доказательств эволюции шеи жирафа не существует.[24] Он приходит к выводу, что «издержки окольного пути — будь то экономические или издержки в виде «заикания» — постепенно увеличивались» по мере развития шеи.[25]
Это правда, что значительная разница в длинах правого/левого ВГН приводит к тому, что импульсы поступают в гортанные мышцы жирафа в несколько разное время, но импульсы вдоль более длинного маршрута левого ВГН настраиваются мозгом так, чтобы мышцы правой/левой гортани функционировали слаженно. Это указывает на предусмотрительность в проектировании, чтобы компенсировать ограничения развития, приводящие к разнице в длине правого/левого ВГН. Хотя жирафы не разговаривают, поэтому не могут заикаться, как утверждает Докинз, и издают очень мало звуков, всё же у них есть функциональная гортань.
Затем Докинз утверждает, что по мере того, как «шея жирафа начала приближаться к своей нынешней впечатляющей длине, общие издержки окольного пути могли бы приблизиться к точке, где — гипотетически — мутантная особь приспособилась бы лучше, если бы у нее нисходящие волокна гортанного нерва отделились от пучка блуждающего нерва и прыгнули через крошечный промежуток к гортани».[26] Он признает, что эта мутация неосуществима, потому что для такой мутации необходимо
«…чтобы произошел своеобразный «прыжок через пропасть», необходимо было бы произвести серьезное изменение — даже переворот — в эмбриональном развитии. Очень вероятно, что необходимая мутация все равно никогда не возникнет. Даже если бы это было так, у неё вполне могли бы быть недостатки — неизбежные при любом крупном перевороте в ходе чувствительного и деликатного процесса. И даже если эти недостатки в конечном итоге могли бы быть перевешены преимуществами обходного пути, предельные издержки каждого миллиметра увеличенного пути по сравнению с существующим невелики. Даже если бы решение «вернуться на исходные позиции» было бы лучшей идеей, если бы его можно было достичь, конкурирующая альтернатива была бы всего лишь крошечным увеличением по сравнению с существующим обходом, и предельные издержки его крошечного увеличения были бы невелики. Я предполагаю, что это меньше, чем издержки «большого переворота», необходимого для того, чтобы получить более элегантное решение».[26]
Этот довод Докинза является прекрасным и обоснованным аргументом против всей теории эволюции путем мутаций. А заключает он тем, что его основная мысль в том, что «возвратный гортанный нерв любого млекопитающего является хорошим доказательством против создателя», ведь
«…это именно то, чего мы ожидаем от эволюции путем естественного отбора, и именно то, чего мы не ожидаем от любого разумного конструктора … если бы это было разработано, никто не мог бы всерьез отрицать, что конструктор допустил серьезную ошибку. Но, как и в случае с возвратным гортанным нервом, все становится ясно, если мы смотрим на эволюционную историю».[26]
Выводы
Левый возвратный гортанный нерв не так плохо спроектирован, как утверждают дарвинисты, а скорее свидетельствует о хорошем и разумном замысле. Не существует никаких доказательств того, что его конструкция вызывает какие-либо вредные последствия, и существует много доказательств в пользу вывода о том, что существующая конструкция является результатом ограничений развития и также служит для тонкой настройки функций гортани. Аргументы, приводимые эволюционистами, являются неверными и препятствуют исследованию конкретных причин существующей конструкции.
Ограничения, возникающие в результате развития плода, на самом деле аналогичны аргументам эволюционистов. Единственное различие заключается в том, что здесь задействованы два различных развития: 1) онтогенез, о котором говорится в этой статье, и 2) филогенез, о котором говорят эволюционисты. Эволюционное «доказательство» становится бесполезным, когда существует столь же обоснованное объяснение, основанное на историческом развитии индивида, его онтогенезе.
Утверждать, что ВГН плохо спроектирован, значит подразумевать, что Богу следовало использовать разные траектории развития эмбриона для всех задействованных структур, чтобы избежать образования петли ВГН вокруг аорты. Тот, кто утверждает, что ВГН плохо сконструирован, предполагает, что существует лучшая конструкция, утверждение, которое не может быть принято, если не будет предложен альтернативный дизайн эмбрионального развития от оплодотворенной яйцеклетки до зрелого плода (включая все неисчислимые молекулярные градиенты, триггеры, генетические каскады, а также анатомические изгибы и складки), который мог бы документально зафиксировать существование улучшенной конструкции. В отсутствие этой информации утверждение о «плохом дизайне» является объяснением «эволюции пробелов». Каждый альтернативный эмбриональный дизайн или путь развития, скорее всего, приведет к своему собственному уникальному набору ограничений, точно так же создавая ложное впечатление плохого дизайна.
Благодарность
Виж Содера (Vij Sodera), Дж. У. Джонс (J.Y. Jones), Клиффорд Лилло (Clifford Lillo), Мэри Энн Стюарт (Mary Ann Stewart), и Джанель Кларк (Janella Clarke).
Ссылки и примечания
1. Prothero, D., Evolution: What the Fossils say and Why it Matters, Columbia University Press, New York, pp. 37–38, 2008.
2. Walsh, F.B., Hoyt, W.F. and Miller, N.R., Anatomy and Physiology of the Optic Chiasm; in: Walsh and Hoyt’s Clinical neuro-ophthalmology, vol. 3, Walsh, F.B., Hoyt, W.F. and Miller, N.R. (Eds.),Williams & Wilkins, Baltimore, MD, pp. 60–69, 1987.
3. Dawkins, R., The Greatest Show on Earth: The Evidence for Evolution, Free Press, New York, p. 360, 2009.
4. Sadler, T.W., Langman’s Medical Embryology, 11th Edition, Williams & Wilkins, Philadelphia, PA, 2010.
5. Mitchell, B. and Sharma, R., Embryology, Churchill Livingstone Elsevier, Philadelphia, PA, 2009.
6. Blechschmidt, E. and Freeman, B., The Ontogenetic Basis of Human Anatomy: A Biodynamic Approach to Development from Conception to Birth, North Atlantic Books, New York, p. 188, 2004.
7. Blechschmidt and Freeman, ref. 6, p. 108.
8. Sturniolo, G., D’Alia, C., Tonante, A., Gangliano, E., Taranto, F. and Schiavo, M.G., The recurrent laryngeal nerve related to thyroid surgery, The American Journal of Surgery 177:487–488, June 1999.
9. Steinberg, J.L., Khane, G.J., Fernanades, C.M.C. and Nel, J.P., Anatomy of the recurrent laryngeal nerve: A redescription, The Journal of Laryngology and Otology 100:919, August 1986.
10. Seikel, J.A., King, D.W. and Drumright, D.G., Anatomy and Physiology for Speech, Language and Hearing, 3rd Edition, Thomson Delmar Learning, New York, p. 186, 2005.
11. Sadler, T.W., Langman’s Medical Embryology, 6th Edition, Williams & Wilkins, Philadelphia, PA, p. 211, 1990.
12. Schoenwolf, G.C., Bleyl, S.B., Brauer, P.R. and Francis-West, P.H., Larsen’s Human Embryology, Churchill Livingstone, Philadelphia, PA, p. 407, 2009.
13. Blechschmidt and Freeman, ref. 6, p. 91.
14. Sadler, ref. 11, p. 212.
15. Sadler, ref. 11, p. 229.
16. Sturniolo et al., ref. 8, p. 487.
17. Armstrong, W.G. and Hinton, J.W., Multiple divisions of the recurrent laryngeal nerve, AMA Archives of Surgery 62(4):532–539, 1951.
18. Sanders, I. and Mu, L., Anatomy of the human internal superior laryngeal nerve, The Anatomical Record 252:646–656, 1998.
19. Bhatnagar, S.C. Neuroscience for the Study of Communicative Disorders. Wolters Kluwer/Lippincott Williams and Wilkins, p. 336, 2008.
20. Colton, R.H., Casper, J.K., Leonard, R., Thibeault, S. and Kelley, R. Understanding Voice Problems: A Physiological Perspective for Diagnosis and Treatment, 3rd Edition, Lippincott Williams and Wilkins, Philadelphia, PA, 2006.
21. Duffy, J.R., Motor Speech Disorders: Substrates, Differential Diagnosis, and Management, 2nd Edition, Elsevier Mosby, p. 42, 2005.
22. Armstrong and Hinton, ref. 17, p. 539.
23. Dawkins, ref. 3, pp. 361–362.
24. Bergman, J., The giraffe’s neck: another icon of evolution falls, J. Creation (previously TJ) 16(1):120–127, 2002.
25. Dawkins, ref. 3, p. 363.
26. Dawkins, ref. 3, pp. 363–364.
Если вам понравилась статья, поделитесь ею со своими друзьями в соц. сетях!
ВАМ БУДУТ ИНТЕРЕСНЫ ЭТИ СТАТЬИ:







