
Примеры теории «один ген – один признак» опровергнуты
Автор: Джерри Бергман
Источник: creation.com
Перевод: Александр Вишняков
Редактура: Владимир Силенок, Катерина Савченко
Я преподавал генетику человека на уровне колледжа и был разочарован, узнав, что в учебнике, которым я пользовался в течение десяти лет, вплоть до 2017[1] года, неверно утверждалось старое, ныне опровергнутое мнение о том, что отдельные распространенные внешние особенности являются результатом экспрессии одного гена. Был исследован ряд заявленных примеров и установлено, что на проявление всех из них влияют несколько генов. Среди этих примеров были отобраны и обобщены десять. Это открытие подтверждает тот факт, что для появления новых признаков, даже кажущихся простыми, требуется целый набор генов, что значительно увеличивает сложность и снижает вероятность появления этих новых признаков путем случайных изменений генотипа, в соответствии с теорией эволюции.
 Ученикам средней школы и студентам колледжей часто преподносят как истину в последней инстанции мысль о том, что многие распространенные человеческие внешние признаки являются примерами простой регуляции одного гена.[2] Это подтверждается распространенными утверждениями, вроде: «Двадцать тысяч лет спустя очередная мутация позволила волосам на нашей голове (в отличие от волос обезьяны или нательных волос человека) расти без ограничений; это – так называемый «ген стрижки»».[3] В учебном пособии по генетике, используемом в колледже, где я преподавал, помимо примеров, обсуждаемых ниже, учитывалась форма лица, кудрявость волос, размер и форма бровей, длина ресниц, способность сворачивать язык и тому подобное, итого, 27 примеров.[1] Исследование, проведенное для подготовки этой статьи, указывает на то, что многие черты, которые долгое время считались результатом одной мутации или экспрессии одного гена, оказались производной экспрессии группы генов.
Ученикам средней школы и студентам колледжей часто преподносят как истину в последней инстанции мысль о том, что многие распространенные человеческие внешние признаки являются примерами простой регуляции одного гена.[2] Это подтверждается распространенными утверждениями, вроде: «Двадцать тысяч лет спустя очередная мутация позволила волосам на нашей голове (в отличие от волос обезьяны или нательных волос человека) расти без ограничений; это – так называемый «ген стрижки»».[3] В учебном пособии по генетике, используемом в колледже, где я преподавал, помимо примеров, обсуждаемых ниже, учитывалась форма лица, кудрявость волос, размер и форма бровей, длина ресниц, способность сворачивать язык и тому подобное, итого, 27 примеров.[1] Исследование, проведенное для подготовки этой статьи, указывает на то, что многие черты, которые долгое время считались результатом одной мутации или экспрессии одного гена, оказались производной экспрессии группы генов.
Изучая влияние факторов, которые приводят к появлению специфических форм носа, ученые недавно обнаружили 14 различных лицевых признаков, проанализировав около 6000 фотографий. Два гена были связаны с формой ноздри (GLI3 и PAX1), один (RUNX2) влиял на форму переносицы, а ген DCHS2, как было обнаружено, влиял на то, насколько далеко нос выпирает наружу, и на угол кончика носа.[4] Без сомнения, существуют и другие гены, влияющие на данные признаки. Макдональд писал: «Тот факт, что в учебниках и лабораторных материалах продолжают сохраняться эти мифы, является позором процесса образования в сфере биологии».[5]
Значение аргумента «один ген — один белок или признак» для эволюции можно легко объяснить на классическом примере часов Уильяма Пейли. Механические часы состоят из сотен частей, каждая из которых функционирует как часть взаимосвязанного устройства. Если какая-либо часть механизма будет удалена, то часы не будут работать должным образом или, скорее всего, не будут работать вовсе. Единственным исключением может являться весь механизм перемещения стрелок, который обычно удерживается четырьмя или шестью болтами, и, если один из них будет удален, то он все еще будет функционировать, но вибрации в том месте, где был удален винт, даже незначительные, могут вызвать небольшую погрешность, которая в конечном итоге может привести к износу и как следствие, менее точному определению времени, что приведёт, в конечном итоге, к поломке.
Точно так же, если большинство генетических функций контролируется несколькими генами, то мутация, даже если она улучшает какой-то аспект организма, скорее всего, окажет неблагоприятное воздействие на другие части живой системы, поскольку, как и в часах, все части организма взаимосвязаны друг с другом, равно как и с другими генами и даже группами генов.
Более того, сама концепция гена теперь размыта из-за неожиданной генетической сложности, проявившейся в том, что дифференциальный сплайсинг интронов может приводить к формированию множества разных белков в результате одной транскрипции гена.[6] Классическое представление о том, что в геноме, якобы, гены-продуценты белков представляют собой отдельные сегменты ДНК, транскрибированные в одну мРНК и в одном направлении, теперь опровергнуто. В настоящее время признано, что несколько генов перекрывают друг друга, занимая одну полосу ДНК, которая производит несколько рабочих мРНК.[7] По оценкам, 94% генов человека производят более одного продукта посредством альтернативного сплайсинга, производя белки с иногда совершенно разными функциями, несмотря на то, что они кодируются одним и тем же геном.[8]
Этот факт утешает тех, кто разочарован небольшим количеством генов в геноме человека: у людей примерно такое же количество генов, как у Caenorhabditis elegans, маленького прозрачного червя. В результате экспресии аналога человеческого гена DSCAM (Downs Syndrome Cell Adhesion Molecule) у фруктовой мушки(Drosophila melanogaster), производится 95 альтернативных сплайсинговых экзонов и еще 38 000 изоформ.[9] Для сравнения, весь геном фруктовой мушки содержит всего 15016 генов. Этот факт позволяет нам возможность увидеть сложную взаимосвязь генома.
Профессор генетики Университета Делавэра Джон Макдональд задокументировал 20 примеров, включая изгиб рук, так называемую «спаржевую мочу» (когда данная жидкость имеет характерный для спаржи запах, а иногда и зелёный цвет), битурия, ямочки на щеках, кудрявость волос и длину пальцев ног, которые обычно используются как примеры истинности постулата «один ген — один признак» на уроках генетики в школах и в колледжах.[10] Был проведен поиск соответствующей литературы по 10 наиболее распространённым примерам концепции «один ген – один признак», отобранным в соответствии с моим опытом преподавания генетики в колледже. Поиск показал, что 10 наиболее распространённых примеров этого мифа контролируются множеством генов.
Примеры опровержения идеи об одном контролирующем гене
Цвет глаз
Типичный пример — это генетическая регуляция цвета глаз, то есть, цвета радужной диафрагмы, которая регулирует количество света, попадающего в глаз. Она выполняет функцию защиты.[11] Например, люди с голубыми глазами часто имеют проблемы со зрением, поскольку отсутствие коричневого пигмента в радужке не полностью защищает сетчатку от неблагоприятного воздействия избыточного количества яркого света.[12] Цвет радужной оболочки в первую очередь зависит от соотношения эумеланина, который дает тёмно-коричневый цвет, и феомеланина, который дает красноватый цвет. Имеет значение так же и распределение меланина по оболочке глаза.[13]
Распространенные цветовые категории радужной оболочки — это голубой, зеленый, карий и коричневый Более подробный анализ указывает на, по крайней мере, девять значений оттенка и насыщенности, а именно: (i) светло-голубой; (ii) синий; (iii) синий с коричневым ободком вокруг зрачка; (iv) зеленый; (v) зеленый с коричневым кольцом вокруг зрачка; (vi) слабый зеленый по краям и коричневый в центре; (vii) коричневый с небольшим вкраплением зелёного по краям; (viii) коричнвый; и (ix) темно-карий.[14] Очевидно, что эти более сложные сочетания требуют гораздо большего количества генетических данных, чем два обычных гена, что и было обнаружено в результате исследования. Кроме того, цвет глаз также может меняться, особенно в раннем детстве.[15] Два основных гена, участвующих в формированиях цветовых вариаций — это гены HERC2 и OCA2, расположенные рядом друг с другом на 15 хромосоме, а также не менее 10 других генов и сложных взаимодействий между ними, влияющие на цвет глаз.[13]
Форма мочки уха
Распространенным упражнением на уроке генетики является изучение типов мочек ушей друг друга и определение количества сросшихся и свободных мочек. Сросшиеся мочки ушей сливаются с боковой стороной головы, а свободные мочки имеют свисающую часть. В большинстве своём у учеников распространены свободные мочки а не сросшиеся.
С точки зрения генетики Менделя, этот признак обусловлен доминантно-рецессивной дихотомией.[16] Они утверждают, что признак свободной мочки является доминантным, а сросшийся – рецессивным, что предполагает, что комбинация генов, контролирующая эти черты, проста.[17] На самом же деле, мочки ушей не делятся на две категории, а точки прикрепления меняются от верхних частей хряща до области за ухом.[18]
Данное наблюдение заставило биологов усомниться в этой упрощенной парадигме еще до появления современной геномики. Еще в 1937 году один анатом предположил, что прикрепление мочки уха может быть полигенным признаком.[19] В одном из новых исследований генетики мочки уха были проанализированы последовательности ДНК и измерения мочки уха у 74 660 человек, включая людей европейского, латиноамериканского и азиатского происхождения. Связывая последовательности ДНК в геноме с моделью развития уха, исследователи идентифицировали 49 областей генов, связанных со способом прикрепления мочки уха. Они также изучили продукты экспрессии генов, активируемых во время развития уха, подтвердив, что различные гены были расположены на многих местах генома. Авторы статьи пришли к следующему выводу: «Эти гены дают представление о сложной биологии развития ушей».[20]
Как и ожидалось, генетика, лежащая в основе этих вариаций, не проста, но она не является приоритетом для исследований, поскольку, насколько известно, вариации мочки уха не связаны с причинами каких-то заболеваний.[21] Вывод таков, что это «миф … будто мочки ушей можно разделить на две четкие категории (свободную и сросшуюся), и что всего один ген с доминирующей аллелью свободной мочки контролирует этот признак».[22] Современные методы исследования теперь подтвердили, что «даже понятие, определяющее ген, размыто совершенно невообразимой и неожиданной сложностью».[23]
Цвет волос
Другим распространенным примером мифа об одном гене является идея о том, будто бы рыжий цвет волос обусловлен единственной рецессивной аллелью с красным цветом. Большинство исследований делят цвет волос на четыре отдельные категории: блонд, рыжий, шатен и брюнет. Как и цвет глаз, цвет волос определяется количеством эумеланина (тёмно-коричневый цвет) и феомеланина (красноватый цвет) в стержне волоса. Количество эумеланина варьируется от очень небольшого, делающего волосы светлыми, до относительно большого количества, при котором образуются чёрные как смоль волосы. Кроме того, у людей с высоким содержанием феомеланина волосы рыжие — от бледно-красного (так называемого «клубничного блонда») до огненно-рыжего и красновато-коричневого.
Поскольку вариации уровня освещённости и различия в цветовых тонах могут влиять на восприятие зрителями цвета волос, Рид использовал рефлекторный спектрофотометр для измерения количества света на разных длинах волн, отражаемого волосами у людей, помеченных как рыжие.[25] Он не обнаружил свидетельств четкого разделения волос на две категории. Вместо этого были отмечены промежуточные цвета, которые нелегко было разделить на красный или не красный. Три наиболее распространенных полиморфизма аминокислот, связанных с рыжими волосами, — это R151C, R160W и D294H, что указывает на то, что в формирование других оттенков волос вовлечено гораздо больше генов.[26]
Цвет кожи
Исследования в области генетики человека опровергли концепцию, в соответствии с которой цвет кожи контролируется лишь несколькими основными генами.[27] В несколько исследований были вовлечены люди с континента с самым большим количеством разнообразия цвета кожи в мире, а именно из Африки. Одно исследование показало, что шесть основных генов вносят только 30% общей изменчивости цвета кожи,[28] а множество других генов ответственны за остальные 70%. В другом исследовании ученые установили, что в общей сложности, есть 15 различных генов, которые вносят основной вклад в цвет кожи.[29]
Палец автостопщика
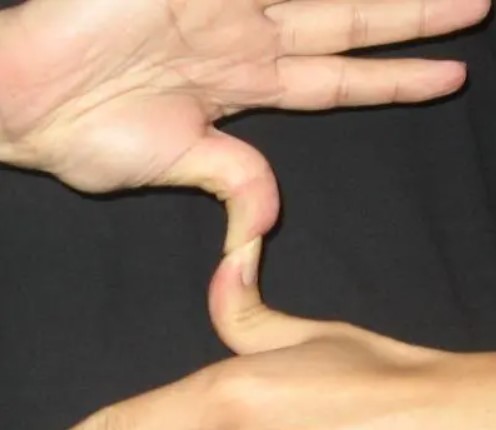
Рисунок 1. Палец автостопщика
«Большой палец автостопщика» — так называют способность сильно согнуть большой палец назад, чтобы образовать большой угол между двумя сегментами кости (рис. 1). Считается, что есть два типа больших пальцев: прямые, когда их нельзя согнуть назад (доминантные), и «большие пальцы автостопщиков» (рецессивные), и этот признак контролируется одним геном с двумя аллелями. Эта идея была предложена Глассом и Кистлером[30] в 1953 году, и с тех пор учителя неоднократно повторяют это утверждение, которое можно было легко подтвердить в большом классе, полном учеников во время урока.
По крайней мере два исследования опровергли этот миф. Харрис и Джозеф[31] использовали рентгеновские снимки 294 человек, чтобы точно измерить угол между первой и второй костями большого пальца. Их анализ показал непрерывное распределение вариаций и указал на то, что у большинства людей были промежуточные результаты, а не только два, как утверждает миф. В аналогичном исследовании с использованием транспортира, удерживаемого на внешней стороне большого пальца для измерения угла наклона, была получена обычная Гауссова кривая.[32] Несмотря на то, что большой палец автостопщика часто используется для демонстрации менделевской генетики, полученные данные опровергают это утверждение: большие пальцы не делятся на две отдельные категории и этот признак не контролируется одним геном.
Вкус Фенилтиокарбамида
Небольшие количества соединений фенилтиокарбамида (PTC) или пропилтиоурацила (PTU), которыми были пропитаны полоски бумаги, пробовались студентами, чтобы определить, является ли вкус этого вещества горьким или наоборот, чувствуется вкус одной лишь бумаги (у тех, кто не чувствителен). Я использовал этот тест с сотнями учеников, преподавая самые разные естественные науки. Идея предложена химиком из компании «Дюпон» (Du Pont) А.Л. Фоксом, работавшим с фенилтиокарбамидом, когда его коллега пожаловался на горький вкус данного химического соединения.[33] Фокс настаивал на том, что это вещество безвкусно, поэтому он попросил своих коллег попробовать PTC, после чего обнаружил, что для некоторых людей он имеет неприятный запах и сильный горький вкус, другие же, как и сам доктор Фокс, сочли фенилтиокарбамид безвкусным.[34]
Гуо и Рид исследовали вопрос вкуса PTC, ссылаясь на 392 ссылки. Они пришли к выводу, что способ, который используется при тестировании, оказывает огромное влияние на результат.[35] В ранних исследованиях кристаллы PTC помещались непосредственно на язык, в то время как в других использовались растворы PTC или бумага, пропитанная PTC, которую затем сушили.[36] Когда этот фактор учитывался, то оказалось, что одни и те же люди, при использовании одного способа оказывались чувствительными ко вкусу, но при использовании другого – нет.[37] Генетический анализ обнаружил связь с полиморфизмами в ДНК в 26 больших семьях, и и вариативность восприятия вкуса была связана с 7 хромосомой и вариациями в хромосоме 16, что указывает на вовлеченность нескольких генов.[38]
Распространенным мифом является утверждение, что существуют только два типа людей – те, кто чувствуют вкус PTC, и те, кто нет. Миф заключается в том, что чувствительность вкуса контролируется одним геном, который доминирует над аллелью, отвечающую за отсутствие вкуса. Дело в том, что «по прошествии почти 70 лет причина этой изменчивости, по-видимому, из-за большого количества генетических факторов, остается загадкой».[39]
Способность свернуть язык в трубочку

Рисунок 2. Пример свертывания языка в трубочку
Некоторые люди могут легко свернуть язык в маленькую трубочку (рис. 2), а другие — нет. Это один из самых распространенных признаков, который преподаватели биологии используют для демонстрации основных генетических принципов, которые можно легко проверить. Этот признак был описан пионером в области генетики дрозофил, Альфредом Стертевантом, описавшим сворачивание языка как простой двух-аллельный признак с доминантной аллелью, отвечающей за способность свернуть язык и рецессивной – за не способность свернуть.[40]
В настоящее время установлено, что навык сворачивания языка в трубочку часто является не генетическим, а приобретённым. Тот факт, что некоторые люди могут научиться сворачивать язык после первой неудачной попытки, свидетельствует о том, что этот признак не является результатом простого генетического воздействия, как считалось ранее. Дополнительным доказательством того, что этот признак не является генетическим, является обнаружение того, что способность скрутить язык может быть разной у однояйцевых близнецов.[41] Доля людей, которые способны сворачивать язык, колеблется от 65 до 81%, причём женщин, могущих это делать, немножко больше мужчин.[42]
Многие овладели этим навыком частично. Некоторые люди могут лишь слегка согнуть кончики языка, потому их нельзя отнести ни к тем, кто умеет, ни к тем, кто не умеет.[43] Несмотря на то, что многочисленные исследования показали, что это умение является не генетическим, а в значительной степени приобретенным, закручивание языка в трубочку остаётся популярным тестом менделевской генетики.[44] В общем, миф о закатывании языка был развенчан.
Вдовий мыс

Рисунок 3. V-обраное место в центре лобной части роста вполос, известное как вдовий мыс.
У некоторых людей есть выступающее V-образное место на передней границе лба и роста волос, называемое вдовьим мысом (рис. 3). Говорят, что аллель «мыса вдовы» доминирует над аллелью «прямой линии» роста волос. Основной проблемой, связанной с этим утверждением, является сложность определения того, у кого есть вдовий мыс, а у кого нет. Есть различные формы границы волос на лбу, потому обычно делается неточное суждение. Одно исследование, проведенное среди студентов-медиков, показало, что только 32 из 1039 человек, что составляет 3%, обладали «небольшим, но заметным» мысом вдовы.[45] Другое исследование с участием 360 женщин показало, что у 81% женщин был мыс вдовы.[46]
Также следует учитывать проблематику возраста. Линия роста волос у многих мужчин и некоторых женщин со временем убывает, часто медленнее посередине, создавая мыс вдовы. Таким образом, довольно сложно отличить залысины от мыса вдовы у взрослых мужчин. Проблема примера мыса вдовы, используемого для иллюстрации основ генетики, заключается в мифе, будто она контролируется одним геном с двумя аллелями, но исследователи так и не смогли определить местонахождение этих генов. Макдональд заявляет, что он не знает, как возник миф, а также нет никаких экспериментальных доказательств, подтверждающих его, несмотря на тщательный поиск в научной литературе.[47]
Сложение рук
Большинство людей предпочитают складывать руки либо большим пальцем левой руки, либо большим пальцем правой руки сверху. Человеку, складывающему руки определенным образом, кажется очень неестественным другой способ. В одном исследовании примерно половина исследуемых людей имели большой палец правой руки сверху, а другая половина — большой палец левой руки сверху.[48] С момента проведения первого более века назад, не было найдено никаких ясных доказательств, подтверждающих, что эти черты соответствуют доминантно-рецессивному мифу.[49]
Обзор почти 100 публикаций, в которых изучался способ складывания рук у людей по всему миру, показал, что у большинства — от 40 до 75% большой палец остается наверху, и только 1% составили те, кому все-равно.[50] В соответствии с одной гипотезой, левши, когда складывают руки, склонны класть большой палец левой руки сверху, а правши большой палец правой руки сверху.[51] В соответствии с другой гипотезой, предпочтения, вероятно, выбираются каждым человеком в раннем детстве и подкрепляются в течение многих лет, до тех пор, пока другое положение станет неестественным. Нет доказательств того, что этот признак является генетическим.
Раздвоенный подбородок

Рисунок 4. Молодой мужчина с раздвоенным подбородком
У некоторых людей есть заметная ямочка или складка на передней части подбородка, называемая расщелиной подбородка (рис. 4). Утверждается, что этот признак контролируется одним геном с двумя аллелями: одна является доминантной и отвечает за расщелину подбородка, а другая является рецессивной, определяющей гладкий подбородок. Немногочисленные доступные генетические данные не подтверждают это утверждение.[52] Основная проблема состоит в том, что многие подбородки находятся между теми, что с расщелиной и гладкими. Подбородки бывают самых разных форм, включая круглые, с ямочками, вертикальные и с Y-образными бороздами.[53] Более того, с возрастом происходит значительное увеличение расщелины подбородка; у около 5% мальчиков в возрасте от 6 до 10 лет есть расщелины подбородка, в то время как уже у 10% мужчин после 35 лет наблюдается расщелина в подбородке. Это изменение с возрастом также свидетельствует против простой генетической модели. Прибавка и потеря веса также влияют на форму расщелины подбородка.
Итог
Чрезмерно упрощенная эволюционная парадигма, которую эволюционисты используют для оправдания своего мировоззрения, не подтверждается исследованиями генома человека, которые последовательно демонстрируют гораздо более высокие уровни сложности в создании этих и других признаков, чем предполагалось ранее. На самом деле, очень немногие признаки являются результатом пары генов, и по мере того, как становится доступным все больше генетической информации, увеличивается и количество различных генов, определяющих большинство признаков. На первый взгляд простые признаки оказываются очень сложными из-за взаимосвязи генетической сети, функционирующей в сложных динамических системах по всему геному.
Применив результаты этого обзора теории «один ген – один белок» к исследованию Гершони и Пьетроковски 2017 года, в котором было идентифицировано 6500 генов, вызывающих половой диморфизм человека (и, следовательно, у мужчин и женщин эти гены выражаются по-разному), результаты не являются неожиданными. Многие признаки, определяющие половой диморфизм, долгое время считавшиеся соответствующими правилу «один ген — один признак», на самом деле не следуют ему.[54] Следовательно, 20 основных признаков, которые, как считается, составляют половой диморфизм человека, на самом деле могут быть определены несколькими сотнями генов, а не двадцатью. Другой фактор — генетическая плейотропия — ситуация, когда один ген влияет на два или более, а часто и на многие другие, которые, на первый взгляд, и не связаны, признаки фенотипа.[55]
Ссылки и примечания
1. Lewis, R., Human Genetics, McGraw-Hill Higher Education, New York, 12th edn, 2017. See Workbook section titled: Observing human phenotypes, pp. 129–139, 2017.
2. Tomkins, J., Human traits: not so simple after all, Acts & Facts 47(2):15, February 2018.
3. Kean, S., The Violinist’s Thumb: And other lost tales of love, war, and genius, as written by our genetic code, Little, Brown and Company, New York, p. 221, 2012.
4. Kaustubh, A. et al., A genome-wide association scan implicates DCHS2, RUNX2, GLI3, PAX1 and EDAR in human facial variation, Nature Communications 7: 11616, 2017.
5. McDonald, J., Myths of Human Genetics, Sparky House Publishing, Baltimore, MD, p.1, 2011.
6. Breitbart, R.E., Andreadis, A., and Nadal-Ginard, B., Alternative splicing: a ubiquitous mechanism for the generation of multiple protein isoforms from single genes, Annual Review of Biochemistry 6(1):467–495, 1987.
7. Andrey, R., Wajngurt, D., Park, N., and Zheng, T., Probing genetic overlap among complex human phenotypes, PNAS 104(28):11694–11699, 2007.
8. Wang, E., Sandberg, R., Luo, S., Khrebtukova, I., Zhang, L., Mayr, C., Kingsmore, S.F., Schroth, G.P., and Burge, C.B., Alternative isoform regulation in human tissue transcriptomes. Nature 456: 470–476, 2008.
9. Neves, G., Zucker J., Daly M., and Chess, A., Stochastic yet biased expression of multiple Dscam splice variants by individual cells, Nature Genetics 36(3):240–246, February 2004.
10. McDonald, ref. 5, p.i, 2011.
11. Brues, A.M., Rethinking human pigmentation, American J. Physical Anthropology 43:387–391, 1975.
12. Sturm, R.A. and Larsson, M., Genetics of human iris colour and patterns, Pigment Cells and Melanoma Research 22(5):544–562, 2009.
13. Liu, F. et al., Digital quantification of human eye color highlights genetic association of three new loci, PLOS Genetics 6:e1000934, 2010; Pośpiech, E., Draus-Barini, J., Kupiec, T. et al., Gene-gene interactions contribute to eye color variation in humans, J. Human Genetics 56(6):447–455, 2011.
14. Mackey, D.A., Wilkinson, C.H., Kearns, L.S., and Hewitt, A.W., Classification of iris colour: review and refinement of a classification schema, Clinical and Experimental Ophthalmology 39(5):462–471, 2011.
15. Matheny, A.P. and Dolan, A.B., Changes in eye colour during early childhood: sex and genetic differences, Annuals in Human Biology 2(2):191–196, April 1975.
16. Lai, L.Y.C. and Walsh, R.J., Observations on ear lobe types, Acta Genetica 16: 250–257, 1966.
17. See the Arizona State University website ‘Ask A Biologist’ for the Mendelian claim which is incorrect as documented in this paper; askabiologist.asu.edu/ mendelian-traits-humans, 2019.
18. Dutta, P. and Ganguly, P., Further observations on ear lobe attachment, Acta Genetica et Statistica Medica 15:77–86, 1965.
19. Wiener, A.S., Complications in ear genetics, J. Heredity 28(12):425, 1937.
20. Shaffer, J.R., Lee, M.K., Roosenboom, J. et al., Multiethnic GWAS reveals polygenic architecture of earlobe attachment, The American J. Human Genetics 101(6):913–924, 2017.
21. Dutta and Ganguly, ref. 18, p. 77.
22. McDonald, ref. 5, p. 14.
23. Tomkins, J.P., Gene complexity eludes a simple definition, Acts & Facts 43(6):9, 2014.
24. McDonald, ref. 5, p. 37.
25. Reed, T.E., Red hair colour as a genetical character, Annals of Eugenics 18(1):115–139, January 1953.
26. Box, N.F., Wyeth, J.R., O’Gorman, L.E., Martin, N.G., and Sturm, R.A., Characterization of melanocyte stimulating hormone receptor variant alleles in twins with red hair, Human Molecular Genetics 6(11):1891–1897, October 1997.
27. Crawford, N.G., Kelly, D.E., Hansen, M.E.B. et al., Loci associated with skin pigmentation identified in African populations, Science 358(6365):eaan8433, 2017; Martin, A.R., Lin, M., Granka, J.M. et al., An unexpectedly complex architecture for skin pigmentation in Africans, Cell 171(6):1340–1353, 2017; Tomkins, J.P., Skin color research confirms biblical narrative, Creation Science Update, Posted on ICR.org, 2 November 2017, accessed 4 December 2018.
28. Wiener, ref. 19, pp. 421–422.
29. Martin et al., ref. 27 p. 1340.
30. Glass, B. and Kistler, J.C., Distal hyperextensibility of the thumb, Acta Genetica 4:192–206, 1953.
31. Harris, H. and Joseph, J., Variation in extension of the metacarpo-phalangeal and interphalangeal joints of the thumb, J. Bone and Joint Surgery 31:547–559, 1949.
32. Glass and Kistler, ref. 30, p. 192.
33. Fox, A.L., The relationship between chemical constitution and taste, PNAS 18( 1):115–120, 1932.
34. As reported in Boyd, W.C., Genetics and the Races of Man: An introduction to modern physical anthropology, Little, Brown and Company, Boston, MA, 1950.
35. Guo, S.W. and Reed, D.R., The genetics of phenylthiocarbamide perception, Annals of Human Biology 28(2):111–142, 2001.
36. Guo, S.W., Shen, F.M., Wang, Y.D., and Zheng, C.J., Threshold distributions of phenylthiocarbamide (PTC) in the Chinese population, Annals of the New York Academy of Sciences 855(1):810–812, 1998.
37. Hartmann, G., Application of individual taste difference towards phenyl-thio-carbamide in genetic investigations, Annals of Eugenics 9(2):123–135, 1939; Lawless, H., A comparison of different methods used to assess sensitivity to the taste of phenylthiocarbamide (PTC), Chemical Senses 5(3):247–256, 1980.
38. Drayna, D., Coon, H., Kim, U.K., Elsner, T., Cromer, K., Otterud, B., Baird, L., Peiffer, A.P., and Leppert, M., Genetic analysis of a complex trait in the Utah Genetic Reference Project: a major locus for PTC taste ability on chromosome 7q and a secondary locus on chromosome 16 p, Human Genetics 112(5–6):567–572, 2003.
39. Guo and Reed, ref. 35, p. 111.
40. Sturtevant, A.H., A new inherited character in man, PNAS 26(2):100–102, 1940.
41. Matlock, P., Identical twins discordant in tongue-rolling, J. Heredity 43(1):24, 1952.
42. Lee, J.W., Tongue-folding and tongue-rolling in an American Negro population sample, J. Heredity 46(6):289–291, 1955.
43. Reedy, J.J., Szczes, T., and Downs, T.D., Tongue rolling among twins, J. Heredity 62(2):125–127, March 1971.
44. Brownie, M., Tongue-rolling myth totally ‘debunked’, usatoday.com/story/ news/nation/2015/08/17/tongue-rolling-myth-totally-debunked/31841559, 17 August 2015.
45. Smith, D.W. and Cohen, M.M., Widow’s peak scalp-hair anomaly and its relation to ocular hypertelorism, Lancet 2(7838):1127–1128, 1973.
46. Nusbaum, B.P. and Fuentefria, S., Naturally occurring female hairline patterns, Dermatologic Surgery 35(6):907–913, 2009.
47. McDonald, ref. 5, p. 67.
48. Reiss, M., The genetics of hand-clasping: a review and a familial study, Annals of Human Biolog 26(1):39–48, 1999.
49. Lutz, F.E., The inheritance of the manner of clasping the hands, American Naturalist 42(495):195–196, 1908.
50. Reiss, ref. 48, pp. 39–41.
51. McManus, D. and Bryden, M.P., The genetics of handedness, cerebral dominance, and lateralization, Handbook of Neuropsychology 6(10):115–144; Child neuropsychology—Part 1, Elsevier Science Publications, New York, 1992.
52. Lebow, M.R. and Sawin, P.B., Inheritance of human facial features: a pedigree study involving length of face, prominent ears and chin cleft, J. Heredity 32(4):127–132, 1941.
53. Günther, H., Anomalien und Anomaliekomplexe in der Gegend des ersten Schlundbogens, Zeitschrift für menschliche Vererbungs-und Konstitutionslehre 23:43–52, 1939.
54. Gershoni, M. and Pietrokovski, S., The landscape of sex-differential transcriptome and its consequent selection in human adults, BMC Biology 15(1):7, 2017.
55. Bergman, J., The pleiotropy problem for evolution, CRSQ 46(4):284–289, Spring 2010.
Если вам понравилась статья, поделитесь ею со своими друзьями в соц. сетях!
ВАМ БУДУТ ИНТЕРЕСНЫ ЭТИ СТАТЬИ:


")




