
Объяснение «робастности» людей
Автор: Др. Питер Лайн (англ. Peter Line)
Источник: creation.com
Перевод: Алена Недоступ
Редактура: Анна Тощёва
В этой статье предпринята попытка объяснить «робастность» (Робастность, измеряемая отношением толщины кости к ее длине, в основном определяется биомеханическим стрессом и наличием андрогенов — прим. переводчика) ископаемых останков древних людей. В статье высказывается предположение о ее связи с продолжительными сроками жизни. В качестве основного механизма, вызывающего робастность, предлагаются изменения в развитии, связанные с долголетием и, скорее всего, связанные с особенностями секреции гормонов щитовидной железы, то есть различное количество тироидных гормонов, доступных для роста скелета на определённых стадиях развития у здоровых людей, по сравнению с современными людьми. Пересматривается природа «хоббита» (Homo floresiensis), чтобы найти объяснение небольшого размера некоторых черепов Homo erectus.
***
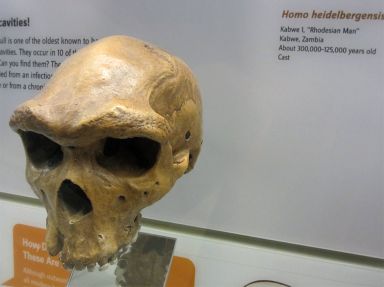
Рисунок 1. Череп (муляж) взрослой особи Homo heidelbergensis Кабве 1 (1325 cc) из центральной Замбии.
Один из интересных вопросов о происхождении человека с позиции креационизма заключается в следующем: почему «робастные» люди, такие как неандертальцы, Homo heidelbergensis и Homo erectus, будучи полностью людьми (то есть потомками Адама и Евы), обычно были более крепкими, грубо сколоченными в своей морфологии, с тяжёлыми и крупными телами или частями тела, по сравнению с более хрупким и субтильным телосложением современных людей с их более тонким и стройным телосложением со слабыми мышечными прикреплениями и костными опорами? Когда речь заходит о «робастной» морфологии ископаемых образцов, таких как Neandertals и H. erectus, часто ссылаются на их черепа, но и остальным костям их скелета также свойственно это качество.
Различия между черепами «робастных» людей
Первый вопрос, который необходимо решить, заключается в том, почему существуют различия между «крепкими» черепами, такими как черепа Neandertals и H. erectus. Как указывал покойный эволюционный антрополог Гарри Шапиро (Harry Shapiro) (почётный председатель Департамента антропологии в Американском музее естественной истории (Department of Anthropology в American Museum of Natural History)), некоторые различия, возможно, были просто связаны с размером: «Рассматривая классический череп Neanderthal, которых сейчас очень много, мы не можем отделаться от убеждения, что его фундаментальное анатомическое строение является увеличенной и развитой версией черепа Homo erectus. Как и у Homo erectus, у него имеется бугристый выступ на затылке, тяжёлый надбровный гребень, относительно уплощённая корона, которая со стороны выглядит как профиль, подобный мансардной крыше. В наиболее широкой точке он низкий, чуть выше ушей, и для него типично отсутствие выступающего подбородка».[1]
Совсем недавно палеоантрополог Даниэль Либерман (Daniel Lieberman), эксперт в так называемой «эволюции» человеческой головы, заметил, что «H. heidelbergensis и H. Neanderthalensis crania в целом похожи на H. erectus, но с небольшим увеличением размера лица и относительного размера мозга. В противоположность им, H. sapiens представляет собой сдвиг в черепно-лицевой архитектуре: с втянутым, меньшим по размеру лицом и более сферическим сводом черепа».[2]
Таким образом, даже с учётом предполагаемых различий между основными группами «робастных» людей, по-видимому, существует ещё большая разница между современными и «робастными» людьми. Рассмотрим «искусственный» вид H. heidelbergensis (рис.1). При осмотре слепков этих ископаемых черепов не бросается в глаза, что именно отличает их от H. erectus, за исключением большего размера некоторых из них. Действительно ли мы имеем дело с разными видами или просто с разными по размеру экземплярами одного и того же вида? Быть может, некоторые из них подверглись различным воздействиям окружающей среды?
По-видимому, общепринятый взгляд, что H. erectus является видом с малым размером мозга, противоречит черепу Йинниушан (Jinniushan), найденному в Китае, который эволюционисты датируют периодом около 200 000 лет назад. Ёмкость этого черепа составляет 1390 см3, и некоторые из них склонны приписать его позднему H. erectus.[3] По–видимому, этот образец легко можно было бы отнести к H. heidelbergensis, в частности из-за того, что он имеет «примитивно-расширенное сочетание» признаков;[3] другие с готовностью причисляют его к H. heidelbergensis.[4] Однако, по мнению некоторых эволюционных экспертов, включение черепа Йинниушан (и некоторых других китайских ископаемых черепов) в группу H. heidelbergensis сталкивается с другими представлениями о происхождении человека, из-за чего это назначение отвергается не из-за морфологии, как таковой, а из-за факторов, связанных с предполагаемым возрастом, местоположением и идеями о расширении мозга.[5] Было высказано интересное предположение, что посткраниальные останки образца Йинниушан напоминают останки неандертальцев.[6] Кажущаяся взаимозаменяемость черепа Йинниушан и H. heidelbergensis, как того требуют соображения удобства, не помогает развенчать идею о том, что H. erectus, H. heidelbergensis и неандертальцы принадлежат к одному и тому же виду. Конечно, могут иметь место региональные различия в форме черепа, как это происходит с разными группами людей сегодня; также не исключено, что черепа, которые особенно малы размером, могут постепенно и непропорционально уменьшаться в некоторых частях. Однако такие различия могут быть вызваны факторами, не имеющими ничего общего с человеческой эволюцией в обычном понимании.[7]
Подумайте о следующем: точно так же, как Вавилонское рассеяние могло привести к тому, что различные группы людей переняли те или иные подмножества генов определённых признаков (например, группы крови), оно также могло привести к тому, что различные группы людей перенимали разные подмножества генов для изменения костных характеристик. Следовательно, по мере того, как группы людей мигрировали из Вавилона, появились варианты «робастных людей», таких как H. erectus, H. heidelbergensis и неандертальцев, возникшие в результате сочетания генетической изменчивости костных признаков и местных факторов окружающей среды, а также установились другие «расовые» характеристики. С течением времени в некоторых группах людей стали доминировать определённые черты (возможно, в результате естественного отбора, но более вероятно, что как результат генетического дрейфа вследствие инбридинга), но те же черты отсутствовали или практически отсутствовали в других группах. Наличие или отсутствие подбородка может быть одной из костных характеристик, которые находятся под генетическим контролем. Большинство «робастных» людей, как правило, имеет неразвитый или отсутствующий подбородок, и это указывает, что такая черта гораздо чаще встречалась в более ранних человеческих популяциях. До сих пор рождаются люди без чётко выраженного подбородка,[8] а значит, это часть нормальной человеческой изменчивости. Однако частота проявления этой черты, безусловно, уменьшилась в сегодняшней популяции.
Долголетие древних людей
Вполне разумно предположить, что большинство этих «робастных» людей жили в первые несколько столетий после потопа, учитывая их общую геологическую связь с пост-потопными отложениями.[9] Таким образом, возникает вопрос: почему древние люди, жившие после потопа, оказались более выносливыми? В предлагаемой нами модели выдвигается предположение, что морфология этих ранних пост-потопных «робастных» людей отражает морфологию их непосредственных предков, живших до потопа, и ответ на вопрос о робастности лежит в долголетии предыдущих поколений людей.
Фактор, который нельзя игнорировать при построении креационистской модели происхождения человека, — это библейская продолжительность жизни отдельных людей в допотопном мире, а также ранняя пост-потопная Земля, информацию о которой мы получаем из Ветхого Завета. В допотопном мире продолжительность жизни, по-видимому, составляла около 900 лет, и даже о людях, рождённых в ранний пост-потопный период (в течение нескольких сотен лет после того, как произошёл потоп), написано, что они жили сотни лет — гораздо больше, чем 70 или около того, как это бывает в наши дни, хотя и сегодня можно найти некоторое количество долгожителей в возрасте более 100 лет.[10]
Долголетие и крепость
Так как же связать продолжительность жизни с робастностью? Если изначально, после грехопадения человека, людям было предназначено жить сотни лет, то это, скорее всего, имеет непосредственное отношение к процессу и срокам развития. Проще говоря, долголетие, вероятно, следует связывать с изменениями в развитии,[11] а не только с процессом старения. С точки зрения замысла, чтобы дольше прожить, возможно, мы изначально должны были быть крепче и сильнее; в таком случае это должно было изначально быть частью процесса развития.
Утолщённые кости свода черепа, грубые черты лица, мощные челюсти и посткраниальные кости, возможно, были необходимы организму, чтобы справиться со столь продолжительным сроком жизни. Рассмотрим также темпы потери костной массы у ныне живущих людей с нормальным процессом старения, когда, начиная с 30-40 лет, «каждое десятилетие женщины теряют около 8% своей скелетной массы», тогда как мужчины теряют 3% за десятилетие.[12] Подобная скорость кажется неприемлемой для продолжительности жизни в сотни лет. Даже в этом отношении что-то должно было отличаться у более ранних людей с большой продолжительностью жизни.
Неандертальцы и Джек Куццо
Креационист Джек Куццо, автор книги о неандертальцах «Похороненный заживо» (Buried Alive, by Jack Cuozzo),[13] давно предположил, что неандертальцы были людьми, жившими после потопа и обладавшими большой продолжительностью жизни. Его позиция заключается в том, что отличительные морфологические признаки неандертальцев являются результатом жизни, достигающей очень преклонного возраста (сотни лет).[14] Например, говорят, что он приписывал форму надбровной дуги неандертальцев тому, что они «сформировались в результате старения и нормального жевания».[15] Он считает, что неандертальские дети демонстрировали более медленную скорость взросления в сравнении с нынешними детьми, и поэтому им требовалось больше времени, чтобы достичь зрелости.[16]
Представленная здесь модель предполагает, что изменения в темпах роста и сроках развития между «робастными» детьми и современными ответственны за различия в «робастности» между взрослыми представителями обеих групп, и что они связаны с генетикой долголетия. Независимо от того, демонстрировали ли неандертальские дети медленную скорость созревания[17] — то есть им требовался более продолжительный период развития для достижения зрелости[18] или завершения роста, —может быть, это так, а быть может и нет, но более длительный период созревания не обязательно следует из изменений в процессах развития, и это не обязательно сопутствует долголетию.
В отличие от Куццо, я считаю, что ключевые особенности морфологии неандертальцев возникли (под влиянием, в той или иной степени, факторов окружающей среды) в процессе развития, а не в процессе старения. Если бы это было связано с процессом старения, то каждый из найденных окаменелых останков неандертальцев должен по умолчанию «прожить» сотни лет, что маловероятно, особенно если некоторые костные характеристики, отличающие этих крепких людей от современных, уже присутствуют в неандертальских образцах, которые, очевидно, все ещё далеки от достижения зрелости, как бы долго это ни длилось.
Однако если характерные для неандертальцев черты были, главным образом, результатом процессов развития (генетически связанных с более высокими факторами долголетия), то некоторые из таких окаменелостей могли принадлежать особям с потенциалом к продолжительной жизни, которые имели грубое строение скелета, но умерли в относительно молодом возрасте. Такое предположение также объясняет наличие этих особенностей у неандертальских детей.
Стоит также отметить, что, по мнению Куццо, образцы, приписываемые H. erectus, следует считать обезьянами.[19],[20] Это противоречит моему убеждению в том, что они были полностью людьми.
Обратите внимание, что «робастные» люди не обязательно жили дольше, как в случае с неандертальцами; многие из них, по-видимому, умирали рано, часто от травм, но обычно не от причин, связанных с преклонным возрастом. По словам Криса Стрингера (палеоантрополога и эксперта по неандертальцам), «они получали множество телесных повреждений», и смерть в возрасте около сорока лет была «очень почтенным возрастом для неандертальцев».[21] Стрингер также отмечает, что: «…проведённое Каспари исследование останков почти семидесяти пяти неандертальцев с места раскопок в г. Крапина (Хорватия) показало, что ни один человек из них, вероятно, не был старше тридцати пяти лет на момент смерти, поэтому вокруг было не так уж много бабушек и дедушек…».[22]
Насколько точны приведенные выше возрастные оценки, сказать трудно. Однако независимо от того, как долго жили отдельные взрослые неандертальцы, они все равно в процессе развития приобретали очень крепкое строение тела, как если бы были генетически запрограммированы жить дольше. И это очень важно, так как именно в процессе развития (а не в процессе старения) появилось большинство отличительных черт морфологии черепа неандертальцев.[23],[24],[25] По словам эволюциониста Пола Джордана (Paul Jordan), «дети неандертальцев интересны тем, что демонстрируют многие из приметных физических отличий их народа, таких как тяжёлые надбровные дуги и общее крепкое строение их тел, из чего можно сделать вывод, что эти признаки рано появлялись в жизни каждого неандертальца».[26]
Тиреоидный гормон
Если бы долголетие было напрямую связано с процессами развития, которые ассоциируются «робастностью», то можно было бы ожидать, что «грубые» черты исчезнут с уменьшением срока жизни. Генетический механизм робастности, вероятно, включает в себя контроль одного или нескольких гормонов, участвующих в росте и поддержании костей,[27] таких как, например, различия в регуляции структуры секреции тиреоидных гормонов между современными и «робастными» людьми. Щитовидная железа вырабатывает тиреоидный гормон, а также гормон тиреокальцитонин. Тиреоидный гормон — это, фактически, два йодсодержащих гормона – тироксин (Т4) и трийодтиронин (Т3), причём Т4 является основным гормоном, секретируемым фолликулами щитовидной железы, а большая часть Т3 образуется в тканях-мишенях путём преобразования Т4 в Т3. Тиреоидный гормон «играет важную роль в регуляции роста и развития тканей. Он имеет решающее значение для нормального развития и созревания скелета и нервной системы, а также для репродуктивных возможностей».[28]
По словам специалиста по эволюции в этой области Сьюзан Крокфорд (Susan Crockford), «отличительная морфология скелета, свойственная неандертальцам, почти наверняка является результатом структуры секреции тироксина (и, следовательно, скорости пренатального и постнатального роста), которая заметно и последовательно отличалась от таковой у современных людей. Неандертальские черты внешне могут напоминать патологические изменения, связанные с врождённым дефицитом йода, поскольку они отражают различное количество тироксина, доступного для роста скелета на определённых стадиях развития, по сравнению со здоровыми современными людьми».[29]
Таким образом, главный переключатель, контролирующий определённый паттерн секреции гормонов щитовидной железы, может существовать у людей с устойчивой морфологией скелета, таких как неандертальцы, что связано с генетическим механизмом долгой жизни. Хотя точный генетический механизм для любого такого паттерна секреции гормонов щитовидной железы (TH) неясен, ключи к разгадке все же есть.
По словам Крокфорд, «в то время как гены в других местах могут изменять конечный ритмический паттерн TH, выход нескольких так называемых «часовых генов», обнаруженных в клетках циркадных осцилляторов, которые находятся в супрахиазматических ядрах (SCN) переднего гипоталамуса, вероятно, является источником пульсирующей продукции… Даже самые незначительные индивидуальные различия в эффективности таких генов (восемь, идентифицированных к настоящему времени) могут иметь драматические последствия для черт, оказывая влияние на секрецию мелатонина – гормона шишковидной железы».[30]
В основном, пульсирующее высвобождение мелатонина из шишковидной железы стимулирует пульсирующую секрецию тиреотропина — высвобождение гормона (TRH) из гипоталамуса, что приводит к всплескам тиреотропного гормона (TSH) из гипофиза, а это, в свою очередь, приводит к пульсирующему высвобождению TH из щитовидной железы.[31]
Крокфорд также утверждает, что «изменения в темпах и сроках пренатального и постнатального развития предков делают возможным большое разнообразие различий в размерах и форме потомственных популяций – хорошо известный эволюционный паттерн, называемый «гетерохронией». Это, без сомнения, самый распространённый способ видообразования… Хотя идентификация точного биологического триггера, который реализует такое изменение в развитии, оказалась труднодостижимой, давно подозревают, что в процесс вовлечены гормоны».[29]
В то время как Крокфорд рассматривает мутации в «наборе генов, которые генерируют фенотипы ритма щитовидной железы… как обеспечение необходимого сырья, индивидуальной вариации, для естественного отбора, чтобы воздействовать на них во время видообразования»,[29] такое эволюционное объяснение не является необходимым.
Гены долголетия
Люди, жившие до и после потопа, вполне могли иметь гены долголетия. Эти гены долголетия, возможно, также вызвали «робастность» или были связаны с генами, вызывающими «робастность».[32] Как вариант, «робастность» могла бы обеспечить долголетие. Однако гены (или генетический механизм), ответственные за это качество жизни, а, следовательно, и за устойчивость, с тех пор были утрачены или деактивированы. Доктор Карл Виланд (Carl Wieland) предположил, что в более ранних человеческих популяциях могли существовать «гены долголетия», которые впоследствии были потеряны в результате генетического дрейфа.[33] По словам Виланда, «исчезновение человеческих линий с более устойчивой морфологией (Neanderthal, erectus) может коррелировать с исчезновением долголетия. «Робастность» может быть результатом генетического долголетия / замедленного созревания или же одни и те же популяции могли иметь [возможно, связанные между собой] гены долголетия и “робастности”».[34]
Изменения в развитии
О характере любых эволюционных изменений в морфологии скелета между «робастными» и современными людьми, как результат возможных различий в характере секреции тиреоидных гормонов, можно только догадываться. Все, что связано с механизмами, регулирующими скорость и сроки развития, очень сложно, и хотя предполагается, что секреция гормонов щитовидной железы является одним из основных факторов, поскольку она играет решающую роль в обеспечении нормального развития и созревания скелета и нервной системы, она не обязательно может быть единственным фактором.
Череп, вероятно, остаётся тем местом, где существуют наиболее существенные различия между «робастными» и современными людьми. Следовательно, ключ к возможным изменениям в развитии можно найти, рассмотрев, как развиваются сохранившиеся человеческие черепа, и что произойдёт, если темпы роста различных частей черепа изменятся. Например, мозг может вносить свой вклад в эти различия. Известно, что человеческое лицо и свод черепа развиваются с разной скоростью, и что это в значительной степени связано с преждевременным или быстрым развитием мозга на ранних стадиях (расширяющийся мозг отвечает за форму свода).[35] Следовательно, если бы развитие мозга замедлилось (или ускорилось), то это могло иметь значительные последствия для формы черепа. Кроме того, само человеческое лицо не растёт равномерно, причём верхняя часть лица[36] изначально растёт быстрее. Вот что говорит об этом эксперт по лицевому развитию Жоффрей Спербер (Geoffrey Sperber):
«Верхняя треть лица первоначально растёт наиболее быстро, в соответствии с ее нейрокраниальными связями и ранним развитием лобных долей головного мозга. Она также первой достигает своего предельного потенциала роста (период интенсивного роста прекращается после 12-летнего возраста). Напротив, средняя и нижняя трети растут медленнее в течение длительного периода, не прекращая роста вплоть до позднего подросткового возраста… Завершение формирования жевательного аппарата прорезыванием третьих моляров (в возрасте от 18 до 25 лет) знаменует прекращение роста нижних двух третей лица».[37]
Сложный характер роста основания черепа также играет важную роль «в определении окончательной формы и размера черепа и, в конечном счёте, морфологии всего черепа, включая окклюзию зубных рядов».[38] Главное в приведённой выше ссылке на рост черепа состоит в том, чтобы подчеркнуть взаимозависимость различных частей черепа (особенности обычно не развиваются изолированно), а также существенную роль, которую рост мозга играет в формировании конечной формы черепа. Следовательно, измените время роста мозга, и вы, скорее всего, измените пространственные отношения между различными частями черепа, что приведёт к различиям в его морфологии».
По мнению Либермана, анатомически современные люди отличаются от неандертальцев и других таксонов Homo «… лишь по нескольким признакам. К ним относятся шаровидная мозговая оболочка, вертикальный лоб, миниатюрная бровь, клыковая ямка и ярко выраженный подбородок. Человек также уникален среди млекопитающих отсутствием лицевой проекции: лицо взрослого H. sapiens почти полностью находится под передней черепной ямкой, в то время как у всех других взрослых млекопитающих, включая неандертальцев, лицо в некоторой степени выступает перед мозговой оболочкой».[39]
У эволюционистов нет проблем с включением изменений в темпах роста для объяснения особенностей предполагаемых ископаемых гоминид. Например, было высказано предположение, что ускорение продольного роста основания черепа на ранних этапах развития неандертальцев, в конечном счёте (из-за более длинного и плоского основания черепа), «повлияло как на форму свода, так и на форму лица и, возможно, было ответственно за многие черепно-лицевые различия между неандертальцами и современными людьми».[40]
Изменения в чертах черепа, как правило, происходят не как самостоятельные события, отдельно от всех других черт, но одновременно с изменениями в других чертах черепа.[41] Например, эволюционисты ссылаются на изменения в форме мозговой оболочки (например, увеличение ширины темени, расширение лобной кости за глазницами и на коронарном шве, увеличение высоты мозговой оболочки и расширение плоскости затылочной кости) как на «побочные продукты расширения коры головного мозга по отношению к стволу мозга, что приводит к увеличению верхней части мозговой оболочки в соотношении с остальными частями черепа».[41]
Некоторые эволюционисты приводят сходные аргументы в отношении лица, предполагая, что «так или иначе» некоторые из наиболее существенных различий в чертах лица («массивные, выделяющиеся надорбитальные выступы, их прогнатические лица и отступающие, лишённые подбородка нижнечелюстные сращения»), которые отличают современные человеческие лица от H. erectus и H. heidelbergensis (цитируемые авторы используют термины Erectines и Heidelbergs), или, более конкретно, утрата этих черт H. sapiens «отражает изменение морфологии зубов».[41] В то время как эволюционисты рассматривают его с точки зрения тенденций человеческой эволюции, эволюционная интерпретация не требуется, поскольку последняя является просто побочным продуктом определённого мировоззрения.
Как насчёт небольшого размера черепов Homo erectus?
Вот мнение эволюционистов Фредерика Кульриджа и Томаса Уинна (Frederick Coolidge, Thomas Wynn): «Размер современного человеческого мозга варьируется от 1100 см3 до почти 2000 см3, и не существует никакой известной корреляции между размером мозга и интеллектом, какой бы способ ни выбирали для его измерения».[42]
Известный учебник по анатомии и физиологии, по существу, говорит то же самое, хотя и расширяет вариации человеческого размера мозга: «Никакой корреляции между размером мозга и интеллектом не существует. Люди с самым маленьким (750 мл) и самым большим (2100 мл) мозгом функционально нормальны».[43]
Несмотря на отмеченную выше огромную вариабельность размеров мозга у ныне живущих людей, которые являются функционально нормальными, возникает справедливый вопрос, почему возможности черепа (коррелирующие с размером мозга) из ископаемых образцов, приписанных H. erectus, в среднем невелики в сравнении с ныне живущими людьми. Некоторые опускаются даже ниже того, что было бы расценено как крайняя граница нормы у ныне живущих людей.[44]
Какие подсказки предлагает «хоббит»?
28 октября 2004 года средства массовой информации по всему миру запестрели заголовками, взятыми из газеты «The Age»: «Утраченная раса человеческих ”хоббитов» обнаружена на индонезийском острове». Обзор первых дней полемики о «хоббитах» был ранее опубликован автором этой статьи.[45] Официально названный Homo floresiensis, образец получил прозвище «хоббит». Первоначальная позиция авторов «Nature paper», объявивших о находке, заключалась в том, что H. floresiensis – новый вид, происходящий от изолированной предковой популяции H. erectus, в которой наблюдается эндемическая карликовость.[46] С тех пор было выдвинуто много гипотез, объясняющих идентичность H. floresiensis, в том числе предположение, что это всего лишь вариант H. erectus,[47] новый вид, происходящий от H. habilis,[48] микроцефальный современный человек,[49] малоразвитый австралопитекоподобный облигатный двуногий[50] и современный человек с кретинизмом.[51]
На мой взгляд, гипотеза кретинизма предлагает наилучшее объяснение посткраниальной анатомии H. floresiensis; существуют убедительные аргументы в пользу того, как с этим согласуется H. floresiensis.[52],[53] По словам палеоантрополога Дина Фалька (Dean Falk), «кретинизм — это состояние замедленного роста и умственной отсталости в результате дефицита гормона щитовидной железы, который может возникать по самым разным причинам, включая питание с недостаточным количеством йода. Велика вероятность рождения младенцев с этим заболеванием у матерей с дефицитом йода. У детей с кретинизмом широкие лица с плоскими носами. Если их не лечить, то с возрастом они становятся меньше ростом, что приводит к карликовости взрослых».
Кретинизм, вызванный дефицитом йода в окружающей среде, не является генетическим нарушением (кретины рождаются у матерей с тяжёлым дефицитом йода),[55] поэтому он может возникать везде, где в пищевой цепи присутствует дефицит йода, и может влиять на людей, живущих в разных частях мира, хотя его клинические особенности могут варьироваться.[56],[57] Дефицит йода и кретинизм оставались проблемой на протяжении довольного долгого периода современной истории, и их до сих пор не удалось победить.[58] Предполагается, что Н. floresiensis страдал от гипотиреоидного эндемического кретинизма (также называемого миксоэдематозным эндемическим кретинизмом); это состояние возникает как результат рождения без функциональной щитовидной железы.[59],[60] Окружающая среда, в которой был обнаружен H. floresiensis, в Лян-Буа на индонезийском острове Флорес, скорее всего, была йод дефицитной.
По мнению эволюциониста Чарльза Окснарда (Charles Oxnard), «сегодняшние показатели разрастания щитовидной железы свидетельствуют о том, что прежние популяции охотников-собирателей в горах должны были испытывать острую нехватку йода и регулярно производить кретинов. Сам Лян-Буа представляет собой известняковую пещеру, а близлежащие почвы являются щелочными и, вероятно, поэтому не содержат йода. Высота над уровнем моря 500 м. Это место достаточно удалено как от северного, так и от южного побережья… Рыбьи кости, найденные в Лян-Буа, — это речные рыбы, которым не хватает йода. Речные воды (река протекает рядом) в таких регионах отличаются низким содержанием йода. Все эти факторы должны были препятствовать доступу к богатым йодом морским продуктам».[61]
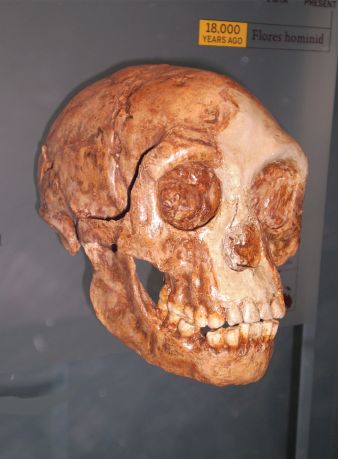
Рисунок 2. Взрослый череп (литой) Homo floresiensis образца LB1 (426 см3) с индонезийского острова Флорес. Является ли он примером «робастного» человека, такого как Homo erectus, страдающего кретинизмом?
Однако все дело осложняет недавний детальный анализ черепа LB1 H. floresiensis (рис. 2), проведённый Юсуке Каифу (Yousuke Kaifu) и др.,[62] включая сравнение с другими черепами, например, H. еrectus из Грузии (Дманиси), череп раннего H. erectus (серии, найденные в Триниле и Сангиране), череп позднего H. erectus (серии, найденные в Самбунгмакане и Нгандонге), черепа китайского H. erectus, ранний и поздний африканские H. erectus, H. heidelbergensis, такие как Кабве и Бодо, а также черепа H. habilis.[63] Хотя они включили в своё исследование большую подборку измерений черепов современных людей, в качестве сравнительной подборки они, по-видимому, не упоминают никаких современных патологических образцов, поэтому нельзя исключить кретинизм или микроцефалию. На основании результатов своих исследований Каифу и его соавторы пришли к выводу, аналогичному тому, что был первоначально предложен в 2004 году: «LB1 наиболее близок к раннему яванскому образцу Homo erectus из Сангирана и Тринила в этих и других аспектах. Мы приходим к выводу, что краниофасциальная морфология LB1 согласуется с гипотезой о том, что H. floresiensis эволюционировал от раннего яванского H. erectus со значительной островной карликовостью».[64]
Соответствует ли череп LB1 модели кретинизма, в частности, небольшим размером мозга? Авторы гипотезы кретинизма утверждают, что, с точки зрения размера мозга (например, пигмейский череп), меньшие родительские популяции могли положить начало кретинизму с эндокринными объёмами менее 500 см3 (территория «хоббита» LB1).[65] В публикации 2010 года они заявили: «Таким образом, были зарегистрированы (см. [7]) и оценены (30) нормальные юго-восточноазиатские пигмоидные черепа объёмом 800-1000 мл. Исходя из этого, на основании измерения роста и объёма мозга среди европейских эндемичных кретинов [7], кретины в этих популяциях могут иметь мозг размером всего 400-500 мл».[66]
Какой сценарий наиболее вероятен? С посткраниальной точки зрения, по-видимому, лучше всего подходит H. floresiensis с кретинизмом, но череп имеет много общих черт и с H. erectus. Вероятный сценарий состоит в том, что H. floresiensis был человеком «робастного» типа (например, H. erectus) с кретинизмом. Ещё раз подчеркнём, что образцы, помеченные как H. erectus, были потомками Адама и Евы и поэтому являются людьми, скорее всего, жившими вскоре после потопа.
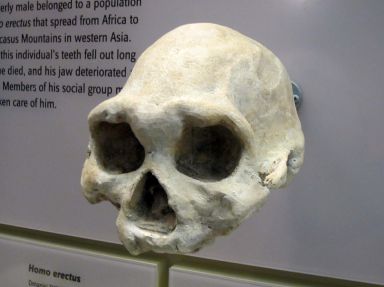
Рисунок 3. «Состаренный» взрослый череп (слепок) Homo erectus образца D3444 (~625 см3) из Дманиси (Грузия). Может быть, это ещё один пример «робастного» человека, страдавшего кретинизмом?
Если череп LB1 H. floresiensis, объем которого оценён в 426 см3,[67] принадлежал патологически робастному человеку с кретинизмом, то это поднимает интересные вопросы о подобной патологии у других слаборазвитых «робастных» людей, таких как образцы H. erectus из Дманиси (рис.3), у четырёх из которых объем черепной коробки колеблется от 600 до 780 см3. Это было бы не так уж удивительно, учитывая, что многие черты кретинизма имитируют так называемые «примитивные» черты эволюции. По словам Окснарда, «примечательно, что так много признаков, сходных с теми, которые обычно присутствуют у больших обезьян, у Australopithecus и Paranthropus, а также у ранних Homo (например, у H. erectus и даже в некоторой степени у H. neanderthalensis), но не у современных H. sapiens, появляются у человека в результате дефицита роста из-за отсутствия гормона щитовидной железы. Другими словами, многие патологические черты кретинизма имитируют примитивные признаки эволюции, что позволяет легко принять патологические черты за примитивные признаки. Распутать этот узел можно лишь путём понимания биологии».[69]
То, что образцы из Дманиси были найдены в одном и том же месте, не так уж необычно. Например, Окснард предполагает, что в «сезонно подвижных группах охотников-собирателей» в прежние времена дети-кретины могли «…во взрослом состоянии подвергаться остракизму со стороны сообщества из-за их ненормальных особенностей и поведения. Будучи не в состоянии легко передвигаться с постоянно переселяющейся общиной, а особенно не в состоянии помочь построить нормальные временные жилища в такой общине, взрослые кретины вполне могли бы отделиться и укрыться в пещерах. Если бы их было разумное количество (по скромным предположениям) 5% от всех рождений, они действительно могли бы укрыться вместе».[70]
Вышеизложенное утверждение, конечно, весьма умозрительно, и возможны альтернативные сценарии, особенно с учётом свидетельств о том, что древние люди заботились о немощных. Может быть, о кретинах заботились, как о группе, более здоровые члены маленького изолированного сообщества.
По данным анализа Каифу и соавторов форма черепа образца LB1 больше всего напоминала череп так называемого «раннего» яванского H. erectus (серия Тринил и Сангиран), а не череп «позднего» яванского H. erectus серии черепов из Нгандонга, также известный как Solo Man. Последние, как правило, имеют большую черепную ёмкость — от 1013 см3 до 1251 см3, тогда как первые черепа (серия Тринил и Сангиран) имеют объем черепа в диапазоне от 813 см3 до 1059 см3.[71] В связи с этим возникает вопрос о том, были ли некоторые из этих мелкочерепных, так называемых ранних яванских H. erectus, также патологически «робастными» людьми с кретинизмом, хотя и менее экстремальным, чем в случае с «хоббитом», или же высокая доля малых размеров мозга у H. erectus является лишь частью нормальных изменений для «робастных» людей?
Если кретинизм по-прежнему остаётся проблемой в современном мире с его развитой медициной и имеющейся в нашем распоряжении информацией о дефиците йода, то насколько большей проблемой он потенциально мог бы быть для ранних пост-потопных/пост-вавилонских человеческих популяций, мигрирующих в неизведанные регионы Земли, скорее всего, ничего не знающих о проблеме (или ее причине) и, вероятно, по горло загруженных работой, просто выживая изо дня в день? Таким образом, популяции «робастных» людей, населяющие любые йододефицитные регионы Африки, Грузии, Китая, Индонезии и т. д., вполне возможно, приобретали высокий уровень заболеваемости кретинизмом.
Вывод
Идеи, изложенные в этой статье, носят теоретический, спекулятивный характер, и, учитывая тематику, вряд ли это может быть иначе. Следовательно, предложенная модель интерпретации появления «робастных» людей, вероятно, нуждается в пересмотре по мере появления большего количества информации по этой теме. Мы надеемся, что изложение этой модели стимулирует дальнейшие дискуссии и исследования в этой области, поскольку она, безусловно, необходима.
Ссылки на источники
- Шапиро Г.Л. Пекинский человек (Shapiro, H.L., Peking Man, Simon and Schuster, New York), стр. 125, 1974.
- Либерман Д.Е. Эволюция человеческой головы (Lieberman, D.E., The Evolution of the Human Head, The Belknap Press of Harvard University Press, Cambridge, MA), стр. 580, 2011.
- Кляйн Р.Г. Карьера человека: биологическое и культурное происхождение человека (Klein, R.G., The Human Career; Human Biological and Cultural Origins, 3rdedn, The University of Chicago Press, Chicago, IL), стр. 348, 2009
- Rightmire, G.P., Brain size and encephalization in Early to Mid-Pleistocene Homo, American J. Physical Anthropology, 124:112, 2004; Tattersall, I., The Fossil Trail: How We Know What We Think We Know about Human Evolution, 2ndedn, Oxford University Press, New York, стр. 281, 288, 2009
- Кляйн, см. п. 3, стр. 312
- Картмилл М., Смит Ф.Х. Родословная человечества (Cartmill, M. and Smith, F.H., The Human Lineage, Wiley-Blackwell, NJ), стр. 431, 2009.
- То есть происхождение от общего предка с дошедшими до нас обезьянами.
- Jacob, T. et al., Pygmoid Australomelanesian Homo sapiensskeletal remains from Liang Bua, Flores: Population affinities and pathological abnormalities, Proceedings of the National Academy of Sciences 103:13423, 2006
- Например, там, где захоронение происходит в пещерах, отложения, внутри которых образовались эти пещеры, часто содержат морские окаменелости после потопа, что указывает на то, что ископаемый человек жил после потопа.
- Wieland, C., Living for 900 years, Creation20(4):10–13, 1998; creation.com/900
- Развитие определяется как «постепенное изменение анатомических структур и физиологических характеристик в период от оплодотворения до зрелости». См.: Мартини Ф.Х., Обер У.К., Нат Дж.Л., Бартоломью И.Ф., Уэлч К. Визуальная анатомия и физиология (Martini, F.H., Ober, W.C., Nath, J.L., Bartholomew, E.F., Garrison, C.W. and Welch, K., Visual Anatomy & Physiology, Benjamin Cummings, San Francisco, CA), стр. 945, 2011.
- Мартини Ф.Х., Нат Дж.Л., Бартоломью И.Ф. Фундаментальные основы анатомии и физиологии (Martini, F.H., Nath, J.L. and Bartholomew, E.F., Fundamentals of Anatomy & Physiology, 9thedn, Pearson Benjamin Cummings, San Francisco, CA), стр. 192, 2012.
- Куццо Дж.У. Погребенный заживо (Cuozzo, J.W., Buried Alive, Master Books, Green Forest, AR, 1998).
- Habermehl, A., Those enigmatic Neanderthals: What are they saying? Are we listening, Answers Research J.3:2,7,8,10, 2010
- Habermehl, см.п. 14, стр. 8
- Habermehl, см.п. 14, стр. 10-12
- Созревание — это процесс достижения зрелости или полного развития. Иногда этот термин используется по-другому, но основное значение (применимое здесь) — это «процесс развития, ведущий к состоянию зрелости». См.: Ребер А.С. Психологический словарь (Reber, A.S., The Penguin Dictionary of Psychology, Penguin Books, London), стр. 422, 1985.
- Зрелость определяется как «состояние полного развития или завершённого роста». См.: Мартини и др., п.12.
- Habermehl, см.п. 14, стр. 3
- Куццо, см. п. 13, стр.101.
- Стрингер К. Происхождение наших видов (Stringer, C., The Origin of Our Species, Allen Lane an imprint of Penguin Books, London), стр. 148–149, 2011.
- Стрингер, см. п.21, стр.152.
- Таттерсаль И. Хозяева планеты: поиски источника появления человека (Tattersal, I., Masters of the Planet; the Search for Our Human Origins, Palgrave Macmillan, New York), стр. 164, 2012.
- Уивер Т.Д. Значение морфологии скелета неандертальцев (Weaver, T.D., The meaning of Neandertal skeletal morphology, Proceedings of the National Academy of Sciences106:16030), 2009.
- Конрой Дж.К. Реконструкция происхождения человека (Conroy, G.C., Reconstructing Human Origins, 2ndedn, W.W. Norton, New York), стр. 524, 2005.
- Джондан П. Неандертальцы: неандертальский человек и история происхождения человечества (Jordan, P., Neanderthal: Neanderthal Man and the Story of Human Origins, Sutton Publishing, Phoenix Mill), стр. 46, 1999.
- Для получения списка гормонов, участвующих в росте и поддержании костной ткани, см. книгу Мартини и Со. (п.12, стр.185).
- Мариеб Э.Н., Хоэн К. Психология и анатомия человека (Marieb, E.N. and Hoehn, K., Human Anatomy & Physiology, 8thedn, Pearson Benjamin Cummings, San Francisco, CA), стр. 609, 2010.
- Crockford, S.J., Commentary: Thyroid hormone in Neandertal evolution: A natural or pathological role? Geographical Review92(1):73–88, 2002.
- Crockford, S.J., Thyroid rhythm phenotypes and hominid evolution: a new paradigm implicates pulsatile hormone secretion in speciation and adaptation changes, Comparative Biochemistry and Physiology Part A135:109, 2003. Иллюстрацию того, как тиоцианаты (SCN) регулируют выброс мелатонина см.: Wright, K., Times of our lives, Scientific American 287(3):43, 2002. Ещё более важным может быть обнаружение прямых нервных связей между SCN и щитовидной железой (по существу, в обход контроля высвобождения TH гипофизом TSH), поскольку это «обеспечивает механизм, который объясняет, каким образом ритмы щитовидной железы могут осуществлять кардиостимулирующий контроль над всеми другими гормонами». Крокфорд С.Дж. Ритмы жизни: тироидный гормон и происхождение видов (Crockford, S.J., Rhythms of Life: Thyroid Hormone and the Origin of Species, Trafford Publishing, Victoria, BC), стр. 88, 2006.
- Крокфорд, см. п.30, стр.107.
- Конечно, если бы существовала связь между генами долголетия и «робастности», и эта связь была бы нарушена, то теоретически мы могли бы получить «робастных» людей с короткой продолжительностью жизни.
- Wieland, C., Decreased lifespans: have we been looking in the right place?, Creation (TJ)8(2):138–141, 1994; creation.com/lifespan.
- Wieland, см. п.33, стр.141.
- Спербер Г.Х. Краниофасциальное развитие (Sperber, G.H., Craniofacial Development, BC Decker Inc., Hamilton, Ontario), стр. 85, 2001.
- Определяется здесь следующим образом: «Верхняя треть лица имеет преимущественно нейрокраниальный состав, причём лобная кость свода черепа, в первую очередь, отвечает за лоб». См.: Спербер, п.35, стр.103.
- Спербер, п.35, стр.103-104.
- Спербер, п.35, стр.100.
- Lieberman, D.E., Sphenoid shortening and the evolution of modern human cranial shape, Nature393:158, 1998.
- Картмилл М., Смит Ф.Х. Родословная человечества (Cartmill, M. and Smith, F.H., The Human Lineage, Wiley-Blackwell, New Jersey), стр. 384, 2009.
- Картмилл и Смит, см. п.40, стр.330.
- Кульридж Ф.Л, Уинн Т. Появление «человека разумного»: эволюция современного мышления (Coolidge, F.L. and Wynn, T., The Rise of Homo sapiens: The Evolution of Modern Thinking, Wiley-Blackwell, West Sussex, UK), стр. 64, 2009.
- Мартини, см. п.12, стр.449.
- Список оценённых черепных способностей Homo erectus и других ископаемых образцов человека, а также других предполагаемых гоминидов см.: Schoenemann, P.T., Hominid brain evolution; in: Begun, D.R. (Ed.), A Companion to Paleoanthropology, Wiley-Blackwell, West Sussex, UK, стр. 142–150, 2013.
- Line, P., The mysterious hobbit, Creation20(3):17–24, 2006.
- Brown, P., Sutikna, T., Morwood, M.J., Soejono, R.P., Jatmiko, Saptomo E.W. and Due, R.A., A new small-bodied hominin from the late Pleistocene of Flores, Indonesia, Nature431:1055, 2004.
- Эта позиция была выдвинута экспертом по Homo erectus Сьюзан Энтон (Susan Anton). См.: Морвуд М., Ван Остерзи П. Новый человек: поразительное открытие и странная история «хоббита» с Флореса, Индонезия (Morwood M. and Van Oosterzee, P., A New Human: The Startling Discovery and Strange Story of the “Hobbits” of Flores, Indonesia, Updated Paperback Edition, Left Coast Press, Walnut Creek, CA), стр. 203, 2009.
- Этой позиции придерживался Колин Гроувз и Австралийского национального (ANU) в Кранберре. См.: Морвуд, п. 47, стр. 203–204
- Микроцефальный человек — это человек с патологией очень маленького размера головы. Именно эта позиция положила начало спору, впервые начатому ныне покойным профессором Теуку Якобом (Teuku Jacob) из Индонезии, также покойным доктором Аланом Торном (Alan Thorne; ранее ANU) и профессором Маси Хеннебергом (Maciej Henneberg, Университет Аделаиды). См.: Лайн, п.45, стр.18-19.
- Хотя первоначально Питер Браун (Peter Brown) предложил идею о карликовом erectus, позже он отказался от неё, предположив вместо этого (в 2009 году), что «гоминиды Лян-Буа прибыли на Флорес в середине Плейстоцена, по существу, со скелетными и зубными характеристиками, которые отличали их, пока они не вымерли около 18 тысяч лет назад. Сравнение с H. erectus, найденным в Дманиси, позволяет предположить, что линия гоминидов Лян-Буа покинула Африку около 1,8 млн лет назад, а, возможно, и до эволюции рода Homo. Мы полагаем, что эти своеобразные, производящие орудия труда, малорослые австралопитекоподобные, облигатные двуногие существа переселились с азиатского материка через Малые Зондские острова на Флорес ещё до появления в этом регионе H. erectus и H. Sapiens». См.: Brown, P. and Maeda, T., Liang Bua Homo floresiensismandibles and mandibular teeth: a contribution to the comparative morphology of a new hominin species, J. Human Evolution 57:592–593, 2009; см. также: Фальк Д. Хроники окаменелостей: каким образом две противоречивые находки изменили наш взгляд на эволюцию человека (Falk, D., The Fossil Chronicles: How Two Controversial Discoveries Changed our View of Human Evolution, University of California Press, Berkeley, CA), стр. 184, 2011.
- Эта гипотеза была впервые предложена в 2008 году в следующей публикации: Obendorf, P.J., Oxnard, C.E. and Kefford, B.J., Are the small human-like fossils found on Flores human endemic cretins?, Proceedings of the Royal Society B275:1287–1296, 2008.
- Oxnard, C., Obendorf, P.J. and Kefford, B.J. Post-cranial skeletons of hypothyroid cretins show a similar anatomical mosaic as Homo floresiensis, PLoS ONE5(9):e13018, 2010.
- Окснард К. Призрачные мускулы, морщинистый мозг, ереси и хоббиты (Oxnard, C., Ghostly Muscles, Wrinkled Brains, Heresies and Hobbits, World Scientific, Singapore), стр. 289–347, 2008.
- Фальк, см. п.50, стр.156.
- Окснард, см. п.53, стр.303, 342.
- Chen, Z-P., Cretinism revisited, Best Practice & Research Clinical Endocrinology & Metabolism24:40–43, 2010.
- Dobson, J.E., The iodine factor in health and evolution, The Geographical Review88(1):4–5, 1998.
- Окснард, см. п.53, стр.333-344.
- Окснард и др., см. п.52, стр.1-2.
- Обендорф и др., см. п.51, стр.1287.
- Окснард, см. п.53, стр.335-337.
- Kaifu, Y., Baba, H., Sutikna, T., Morwood, M.J., Kubo, D., Saptoma, E.W., Jatmiko, Awe, R.D. and Djubiantono, T., Craniofacial morphology of Homo floresiensis: Description, taxonomic affinities, and evolutionary implication, Human Evolution61:644–682, 2011.
- Каифу и др., см. п.62, стр.653.
- Каифу и др., см. п.62, стр.644.
- Окснард и др., см. п.52, стр.7.
- Окснард и др., см. п.65. Обратите внимание, что цитата в п.7 ссылается на Обендорфа (и др.) из п.51. Также заметьте, что в цитате (п.30) ссылка на Berger, L.R., Churchill, S.E., De Klerk, B. and Quinn, R.L., Small-Bodied Humans from Palau, Micronesia, PLoS ONE3(3):e1780, 2008.
- Kubo, D., Kono, R.T. and Kaifu, Y., Brain size of Homo floresiensisand its evolutionary implications, Proceedings of the Royal Society B 280:20130338, 2013; http://dx.doi.org/10.1098/rspb.2013.0338.
- Schoenemann, ref. 44, pp. 144; Lordkipanidze, D. et al, A fourth hominin skull from Dmanisi, Georgia, The Anatomical Record Part A288A:1150, 2006.
- Окснард, см. п.61, стр.342.
- Окснард, см. п.61, стр.339.
- Шенеманн, см. п.44, стр.144-145.
Если вам понравилась статья, поделитесь ею со своими друзьями в соц. сетях!
ВАМ БУДУТ ИНТЕРЕСНЫ ЭТИ СТАТЬИ:







